


Mitochondrial dysfunction in chronic neuroinflammatory diseases (Review)
- Authors:
- Published online on: April 2, 2024 https://doi.org/10.3892/ijmm.2024.5371
- Article Number: 47
-
Copyright: © Qin et al. This is an open access article distributed under the terms of Creative Commons Attribution License.
Abstract
1. Introduction
Neuroinflammation is a response orchestrated by the central nervous system (CNS) in response to infection and injury. The acute neuroinflammatory response reduces damage by promoting the repair of injured tissue. However, persistent stimulation leads to the transformation of the inflammatory response from acute to chronic, resulting in neuronal functional impairment and thus facilitates the progression of CNS diseases (1). Chronic neuroinflammation has been reported as a pathological feature present in several neurodegenerative diseases such as Alzheimer's disease (AD), Parkinson's disease (PD), multiple sclerosis (MS) and epilepsy, amongst other neurological disorders (2,3). The close association between neuroinflammation and neurodegeneration suggests that neuroinflammatory mechanisms may trigger neuronal degeneration, leading to neurotoxicity and a loss of neuronal cells.
Microglia, which are activated by pathological stimuli such as infections, foreign pathogens and neurodegeneration, produce chemotactic factors and proinflammatory cytokines, including nitric oxide, reactive oxygen species (ROS), interleukin (IL)-1β, IL-6 and tumor necrosis factor-α (TNF-α), to eliminate detrimental elements. Nonetheless, persistent stimulation can lead to an overabundance of inflammatory factors, which in turn inflict damage upon neurons (4). Likewise, astrocytes exhibit dual roles in neuroinflammation. In response to cerebral trauma, astrocytes undergo proliferation and transition into a neuroprotective state, fostering reparative and regenerative mechanisms such as remyelination (5). Conversely, in neuroinflammatory diseases, astrocytes are excessively activated into a neurotoxic state by cytokines secreted by microglia, releasing uncontrolled pro-inflammatory cytokines and complement proteins, which exacerbate damage to neighboring cells (6,7). Moreover, astrocytes facilitate lymphocyte movement across the blood-brain barrier, engage in antigen presentation between lymphocytes and microglia, and activate peripheral B cells and T cells via the lymphatic system, thereby amplifying the cerebral inflammatory cascade (1,8,9).
In the CNS, mitochondria serve as the primary energy source for cellular metabolic processes and are pivotal in regulating cellular metabolism, calcium signaling and programmed cell death (10). Neurons rely on mitochondrial oxidative phosphorylation (OXPHOS) to meet their energy demands, maintain ion gradients and facilitate neurotransmitter uptake and recycling (11). Astrocytes contribute to the mitigation of neuronal and oligodendrocyte free fatty acid peroxidation and ROS generation through mitochondrial fatty acid β-oxidation (FAO), which accounts for ~20% of the brain's energy supply (12). Additionally, mitochondrial calcium ions modulate key enzymes of the tricarboxylic acid cycle, such as pyruvate dehydrogenase, thereby regulating OXPHOS (13). Mitochondrial calcium ions also regulate synaptic communication and excitability by modulating astrocytic proliferation and the release of excitotoxic glutamate (14). In addition, microglial mitochondria enhance migration and phagocytosis through calcium ion influx (15). Once mitochondrial dysfunction occurs, the mitochondria become insufficient to meet the heightened energy demands of overstimulated neurons and hyperactive glial cells, leading to abnormal cell metabolism and widespread cell death (10).
Mitochondrial dysfunction serves as both a cause and a consequence of chronic neuroinflammatory diseases. Neuronal mitochondrial dysfunction has been observed in AD, PD and amyotrophic lateral sclerosis (16-18). Chronic inflammation leads to the secretion of cytokines that sustain inflammation and redox stress, inducing mitochondrial DNA (mtDNA) damage (19). Correspondingly, damaged mitochondria can further induce persistent inflammatory responses and downstream pathological inflammation (20,21). A study has shown that inhibiting mitochondrial complex I activates microglia, whereas inhibiting mitochondrial fission reduces pro-inflammatory cytokine generation (12). Due to mitochondrial damage, overactivated microglia undergo a metabolic shift from OXPHOS to glycolysis, resulting in increased generation of ROS and reactive nitrogen species (RNS), thereby exacerbating the inflammatory response (22). Concurrently, microglia can induce the generation of pro-inflammatory astrocytes by releasing fragmented mitochondria (23). Furthermore, impairment of mitochondrial FAO in astrocytes contributes to the development of neuroinflammation and subsequent neurodegenerative processes (24). Additionally, the accumulation of damaged mitochondria in neurons can accelerate the progression of diseases by initiating programmed cell death (23). Mitochondria may therefore be a key link between chronic neuroinflammation and the pathogenesis of neurodegenerative diseases. Thus, repairing mitochondrial dysfunction may improve the outcomes of neurodegenerative diseases, such as AD and PD (25,26).
The aim of the present review was to summarize the molecular characteristics of mitochondrial dysfunction and provide potential directions for targeting mitochondria in the treatment of chronic neuroinflammatory diseases.
2. Pathogenesis of mitochondrial dysfunction
Mitochondrial dysfunction
Mechanisms of mitochondrial dysfunction involved in the progression and prognosis of chronic neuroinflammatory diseases include oxidative stress, epigenetics, mitochondrial dynamics and calcium homeostasis (27-32) (Fig. 1).
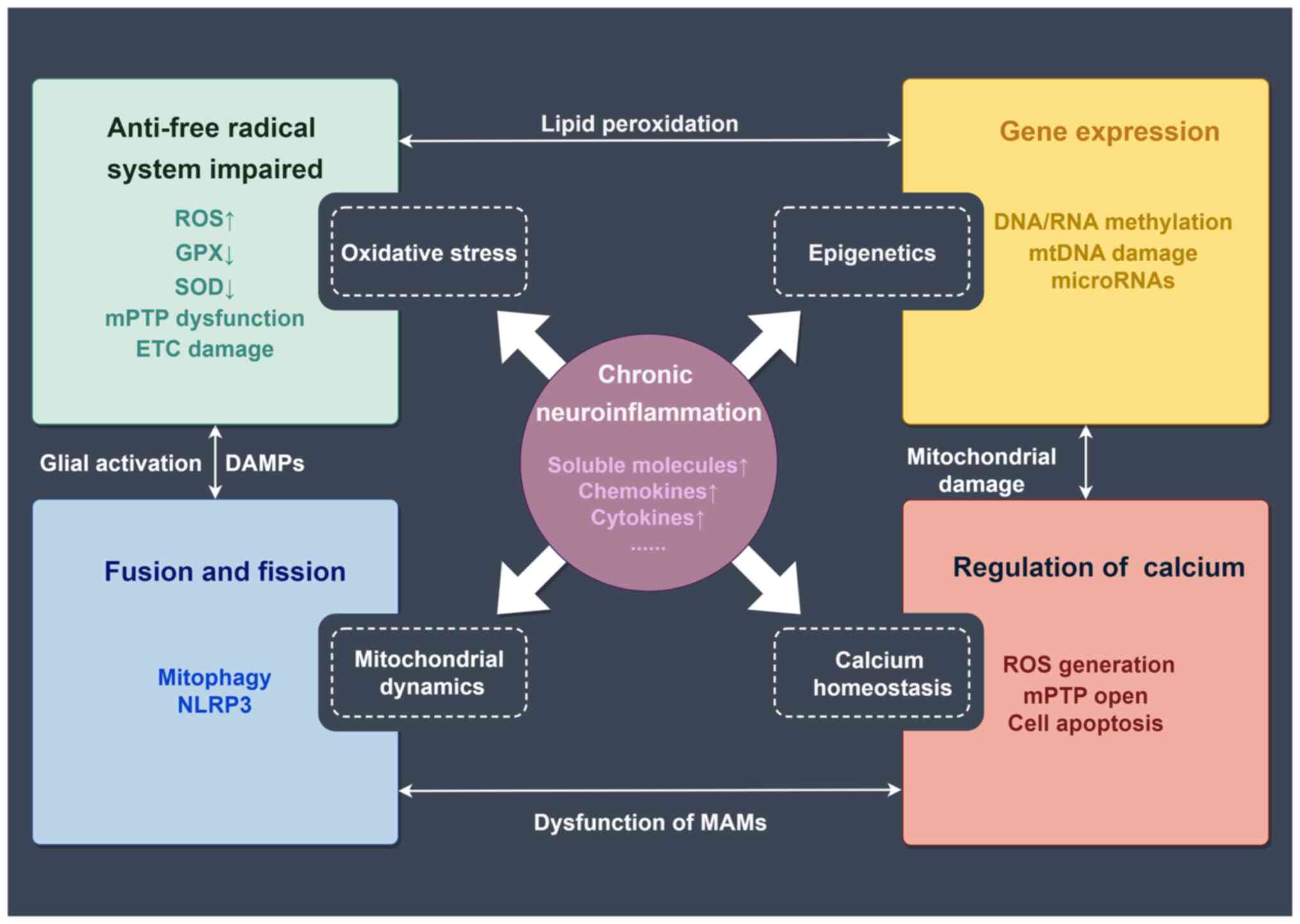
Oxidative stress
In normal, healthy cells, 90% of ROS are generated as a result of cellular respiration. During this process, electrons detach from the electron transport chain and attach to oxygen, producing superoxide anions (O2−) (33). Additionally, metal enzymes present within organisms utilize the interaction between oxygen and metal ions to generate ROS, which is a result of cellular metabolism (34). Conversely, normal cells also possess a protective system against free radicals, primarily composed of antioxidant enzymes such as glutathione peroxidase (GPX), non-enzymatic antioxidant factors, superoxide dismutase (SOD) and catalase (33). The excessive reduction of free radicals is catalyzed by antioxidant enzymes. SOD acts on O2− to produce hydrogen peroxide (H2O2), which has a lower oxidative capacity than O2−, while catalase and GPX enzymes, with the assistance of certain cofactors, convert H2O2 into H2O. When this regulatory process is disrupted, ROS can inflict destructive damage on cells (33). Excessive ROS further induces peroxidation modifications of cellular macromolecules such as lipids, proteins, RNA and DNA (35). For instance, protein peroxidation may acquire toxic functions by forming cytotoxic aggregates. Therefore, the accumulation of ROS caused by various factors (such as increased oxygen consumption in the brain due to high energy demand, elevated levels of unsaturated fatty acids in neuronal membranes, high levels of redox transition metal ions, low antioxidant levels and neurotransmitter oxidation) makes the brain highly susceptible to the damaging effects of oxidative stress (36). The excessive generation of ROS, leading to oxidative stress, has emerged as a shared underlying mechanism implicated in multiple chronic neuroinflammatory disorders, such as AD and PD (37,38).
Due to being the primary source of ROS, mitochondrial dysfunction appears to be a potential focal point for the underlying pathology of neuroinflammation (39). Excessive free radicals damage the inner mitochondrial membrane, leading to compromised mitochondrial energy production and metabolism in the brain. This results in neuronal dysfunction and further exacerbates oxidative stress, promoting neuronal dysfunction and apoptosis. Furthermore, free radicals can directly or indirectly induce abnormal mitochondrial permeability transition pore (mPTP) function, indirectly altering the fluidity, permeability and osmotic properties of the mitochondrial membrane, thereby facilitating mPTP-related ROS release (40). Moreover, free radicals also interfere with electron transport chain (ETC) complexes, further promoting ROS generation (41). A study has also reported that mitochondrial ROS (mtROS) can lead to impairment of complex I within the mitochondrial ETC. This in turn results in a reduction of mitochondrial OXPHOS efficiency (42). This cycle formed by mitochondrial dysfunction and inflammation-related oxidative stress exacerbates the pathological damage in neuroinflammatory disorders.
Mitochondrial epigenetics
Epigenetic modifications within the mitochondria can influence mitochondrial gene expression and function. In neuroinflammatory conditions, heightened ROS production induces deleterious effects on mitochondrial respiration and OXPHOS, leading to DNA oxidation, rearrangements and mutations (43). mtDNA, with its elevated mutation rate and proximity to OXPHOS sites, is more susceptible to oxidative stress compared with nuclear DNA (43). Methylation is a primary epigenetic mechanism within mitochondria due to the absence of histones in mtDNA (44). Decreased mtDNA methylation levels have been observed in blood samples and postmortem brain tissues from individuals with neuroinflammatory diseases (44). Additionally, chronic stress activates the hypothalamic-pituitary-adrenal axis, resulting in excessive glucocorticoid release and the regulation of mtDNA transcription and mtRNA expression (45). Conversely, mtDNA mutations exacerbate ROS production and trigger apoptosis through disruptions in the electron transport chain, impaired protein synthesis, and increased replication errors, influencing disease onset and progression (46). MtDNA damage results in impaired ETC function, reduced ATP generation, increased levels of ROS and disrupted calcium homeostasis, leading to exacerbated amyloid-β (Aβ) processing and aggregation in AD mice (47). Another study reported that Aβ induced mtDNA methylation, which persisted after the removal of Aβ and induced cognitive impairment in AD (48). Furthermore, non-coding RNAs are implicated in chronic low-grade systemic inflammation, known as inflammageing, impacting the energetic, oxidative and inflammatory status of senescent cells by modulating NF-κB/NLR family pyrin domain containing 3 (NLRP3) pathways and triggering senescence-associated secretory phenotype (43). Downregulation of microRNAs could also promote neuroinflammation by affecting the expression of genes critical for neuronal function and immune response in PD (49). It is evident that mitochondrial epigenetics is closely associated with the development of neuroinflammation. Understanding the regulatory role of mitochondrial epigenetics is therefore crucial for unraveling the underlying mechanisms of various neurological disorders.
Mitochondrial dynamics
The complex processes of mitochondrial fusion and fission collectively maintain mitochondrial functionality in the face of cellular metabolic or environmental stress (50). Mitochondrial fusion involves the merging of individual mitochondria, resulting in larger and interconnected networks. This process allows for the exchange of contents, including proteins, lipids and mtDNA, thereby promoting functional complementation and maintaining mitochondrial integrity. Mitochondrial fission is the opposite of fusion and involves the division of mitochondria into smaller fragments. Fission has a crucial role in quality control mechanisms, as it allows for the removal of damaged or dysfunctional portions of mitochondria through a process termed 'mitophagy' (51). Moreover, mitochondrial fission also facilitates the distribution of mitochondria throughout the cell (52).
Chronic neuroinflammation disrupts mitochondrial dynamics. Under neuroinflammatory conditions, dysfunctional mitochondria release ROS and damage-associated molecular patterns (DAMPs) (53), activating microglia and astrocytes, and triggering the release of pro-inflammatory cytokines and chemokines (54), thereby exacerbating the damage. In addition, disruption of mitochondrial dynamics can result in the accumulation of dysfunctional mitochondria, leading to increased susceptibility to inflammation-induced neuronal death. For instance, microglia can activate astrocytes into a neurotoxic state by releasing mitochondrial fragments and damaged mitochondria, further mediating extracellular neuronal death in neuroinflammation (23). Furthermore, mitophagy also proves advantageous in eliminating impaired mitochondria and decreasing the infiltration of inflammatory molecules at the location where damaged mitochondria accumulate (55).
Mitochondrial dynamics may also have a significant involvement in inflammasome activation in chronic inflammation. Inhibition of dynamin-related protein 1 (Drp1) and overexpression of fusion proteins can attenuate inflammation-associated inflammasome responses (56). During RNA virus infection, mitofusin-2 interacts with NLRP3 to activate inflammasomes (57). Thus, molecules involved in mitochondrial dynamics may be crucial regulators of inflammasome activation. In summary, mitochondrial dynamics are essential for maintaining neuronal health and survival and a balanced fusion and fission process is beneficial for maintaining healthy mitochondrial function.
Mitochondrial calcium homeostasis
Mitochondrial calcium homeostasis holds significant importance in maintaining the functionality of neurons and glial cells (58). Mitochondrial function not only sustains the energy prerequisites of both spontaneous and induced neuronal activities in the brain through energy metabolism, but also governs neuronal signaling via uptake and cycling of mitochondrial calcium ions (59). Furthermore, the dynamic regulation of mitochondrial calcium homeostasis is also important for cell survival (13).
In the progression of neuroinflammatory diseases, calcium homeostasis remains a crucial molecular mechanism (60). The dysregulation of neuronal calcium homeostasis leads to oxidative stress, mitochondrial dysfunction, protein conversion disorders and neuroinflammation (61). Activated glial cells serve a crucial role in neuroinflammation and release soluble signaling molecules, including chemokines, pro-inflammatory cytokines, glutamate, prostaglandins, ROS, RNS and damaged mitochondria (62-64). Astroglial calcium signaling appears to be dysregulated in AD, which is potentially linked to the accumulation of Aβ in the brain (65). Depletion of mitochondrial calcium transporters has been shown to mitigate the inflammatory damage caused by glial cells activated by lipopolysaccharides (66).
Mitochondrial Ca2+ accumulation stimulates oxidative metabolism by modulating Ca2+-sensitive dehydrogenases and metabolite carriers (67). Calcium homeostasis disruption can lead to an excessive buildup of matrix Ca2+ and subsequent initiation of the mPTP, thereby affecting mitochondrial function (68). This results in a decrease in ATP synthesis and an increase in ROS generation (69,70). Similar to Ca2+, ROS also serve a crucial role in initiating the opening of the mPTP. This event results in mitochondrial swelling and impairment of the respiratory chain, thereby exacerbating oxidative stress-induced damage (71). Notably, mitochondria-associated membranes (MAMs) are specialized regions that regulate endoplasmic reticulum (ER) contact and transmit Ca2+ into mitochondria. These contact sites facilitate the exchange of various molecules, including lipids and signaling molecules, between the ER and mitochondria (72). Dysregulation of MAMs can therefore affect calcium signaling and disrupt communication between the ER and mitochondria, contributing to the activation of glial cells and the release of pro-inflammatory molecules (73,74). MAMs have also been found to regulate autophagy and mitochondrial dynamics (75). Dysregulation of MAMs has been observed in numerous neuroinflammatory disorders, such as AD and PD (76). These finding imply that targeting MAMs may hold promise as a therapeutic approach for the treatment of neuroinflammatory diseases.
3. Crosstalk of mitochondria in chronic neuroinflammation
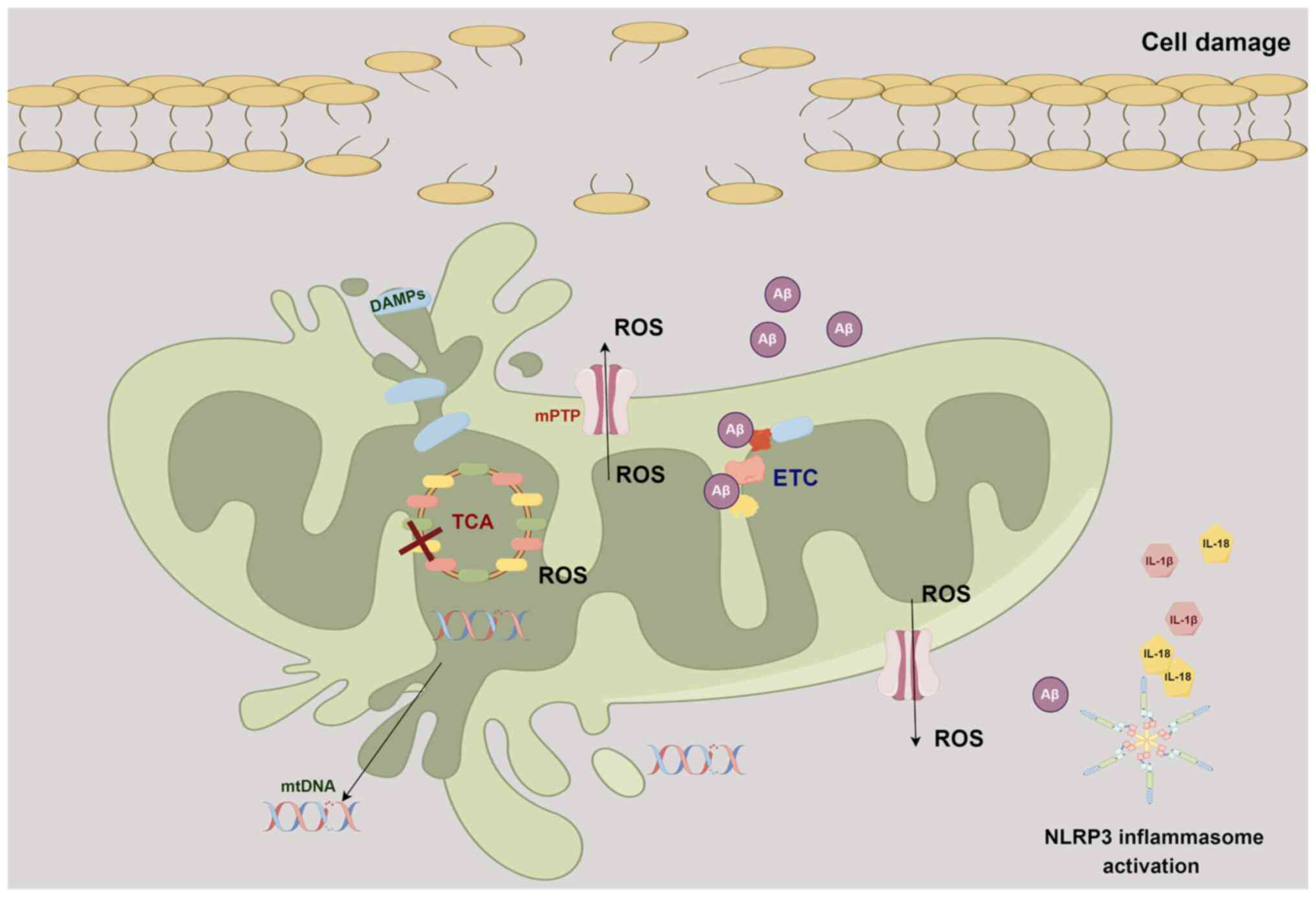
AD
AD is a gradually advancing neurodegenerative condition characterized by the presence of Aβ and τ protein tangles, which are considered distinctive pathological markers. Aβ accumulation has also been observed within the mitochondria in the brains of patients with AD and transgenic AD mouse models (77,78). Aβ can directly disrupt the ETC and interfere with various mitochondrial matrix proteins and putative components of the mPTP, ultimately resulting in mitochondrial dysfunction (77-80).
In the early stages of AD, another often observed mitochondrial abnormality is the excessive generation of ROS, culminating in an upsurge of oxidative stress (81). Oxidative stress causes neuronal cell death, which contributes to the progressive cognitive decline seen in AD (82). Normally, mitochondria serve as pivotal guardians of the cellular redox equilibrium, orchestrating this balance via their antioxidant defense systems. However, malfunctioning mitochondria compromise these protective mechanisms, resulting in diminished scavenging of ROS and an escalation in oxidative harm (83). Furthermore, surplus ROS within mitochondria harms lipids and proteins. For instance, lipid peroxidation engenders the production of harmful byproducts such as malondialdehyde and 4-hydroxynonenal, intensifying the oxidative stress milieu (84). Concomitantly, protein oxidation can induce structural and functional impairments in mitochondrial proteins, thereby impacting energy synthesis and overall integrity. It is also worth noting that the heightened oxidative stress observed in AD can precipitate mutations, deletions and impairments in mtDNA repair mechanisms (85). These events further compound mitochondrial dysfunction, instigating a cycle of oxidative stress and neuronal damage.
Furthermore, DAMPs released from compromised mitochondria, coupled with elevated ROS levels, serve to intensify immune responses, with microglia playing a pivotal regulatory role in this process (86). On the one hand, activated microglia contribute to reducing neuroinflammation by phagocytosing and eliminating Aβ, while on the other hand, these microglia release pro-inflammatory cytokines and other inflammatory molecules, thus promoting inflammation (86,87). Notably, emerging research suggests that there is a bidirectional communication between mitochondria and microglia (88,89). Damaged mitochondria release mtDNA fragments into the cytoplasm, which can activate immune responses through Toll-like receptor 9, NLRP3 and stimulator of interferon genes (STING) signaling pathways. Microglia recognize these mtDNA fragments as danger signals and respond by releasing inflammatory mediators that further amplify the inflammatory microenvironment, inducing mitochondrial damage and subsequent cell death (89). This communication may also perpetuate neuroinflammation and contribute to the progression of AD. In addition, activation of the NLRP3 inflammasome is also an important factor in the pathogenesis of AD (90,91), leading to the release of potent pro-inflammatory cytokines such as its effector molecule, IL-1β (92). Elevated IL-1β levels have been detected in the serum, cerebrospinal fluid and brain tissues of patients with AD (93). IL-1β can enhance the neuronal production of Aβ and induce τ protein phosphorylation, the blocking of which can alleviate neuroinflammation by reducing Aβ levels and τ activation (94,95). IL-18, another proinflammatory cytokine released when the NLRP3 inflammasome is activated, has been demonstrated to be correlated with susceptibility to sporadic late-onset AD (92,96). In conclusion, the neuroinflammatory process plays a crucial role in AD development, and understanding the intricate relationship between mitochondria and neuroinflammation in AD offers potential therapeutic avenues (Fig. 2).
PD
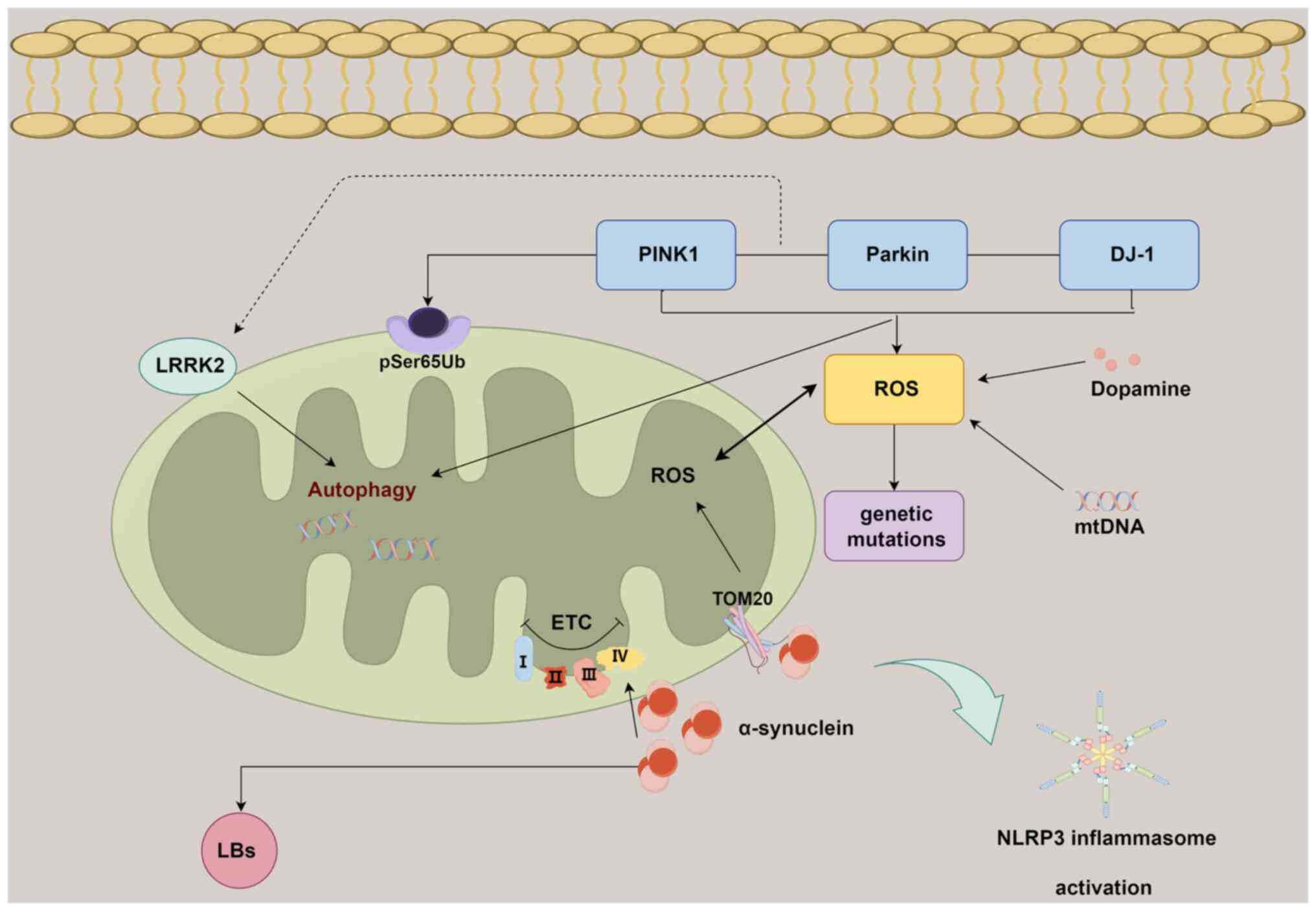
PD is the second most prevalent neurodegenerative disorder, succeeding AD, with a primary impact on the elderly population (97). PD is characterized by the subclinical presence of cytoplasmic proteinaceous aggregates, specifically α-synuclein, which congregate to form Lewy bodies (LBs) within the substantia nigra. This process is coupled with a decline in dopaminergic (DA) neurons. The extensive degeneration of these DA neurons results in diminished dopamine levels within the brain, which gives rise to a spectrum of clinical manifestations, encompassing challenges in maintaining posture, the emergence of stationary tremors, a decrease in movement speed (bradykinesia) and the onset of joint stiffness reminiscent of ankylosing arthritis (49). Following the loss of dopamine-producing neurons, there is a subsequent degeneration of other neuronal subtypes, giving rise to symptoms unresponsive to dopamine modulation. These symptoms encompass a spectrum of manifestations, including insomnia, compromised olfactory perception, dysregulation of the autonomic system, pain perception alterations and sensory dysfunction (98). However, the precise pathophysiological mechanisms driving PD have remained elusive. In addition to α-synuclein, there is mounting evidence implicating other genetic mutations such as in Parkin, leucine-rich repeat kinase 2 (LRRK2) and DJ-1, alongside environmental factors (including exogenous neurotoxins, age and diet) as potential contributors to the etiology of PD (99,100). These factors intricately contribute to the processes of neurodegeneration and neuroinflammation stemming from oxidative stress, α-synuclein oligomerization and mitochondrial dysfunction. Notably, it has been observed that α-synuclein is also present on the mitochondrial surface, exerting an impact on mitochondrial structural integrity and functional dynamics (101). Impaired complex I has been found in samples from patients with PD and the introduction of toxins inhibiting complex I has been shown to lead to the loss of dopaminergic cells and the manifestation of Parkinson's disease symptoms (102). Furthermore, the presence of mutations in mtDNA has been identified within neurons of individuals with PD (103). As a result, mitochondrial dysfunction emerges as a recurring determinant in the context of PD.
The PTEN-induced kinase 1 (PINK1)/Parkin pathway serves a pivotal role in the context of mitochondrial dysfunction and its association with PD. In the current understanding of PD, mutations within the PINK1 (PARK6) and Parkin (PARK2) genes are thought to be associated with the manifestation of autosomal recessive early-onset PD (104). Research has demonstrated that the PINK1/Parkin pathway participates in the progression of PD by influencing mitochondrial autophagy (102,103,105). While mice lacking PINK1 or Parkin do not exhibit significant PD-related phenotypes, a study has demonstrated that these mice accumulate mtDNA mutations, which consequently promotes inflammation in aged Parkin−/− (also termed 'Mutator') mice. This pathological progression appears to be modulated by STING signaling (106). Furthermore, elevated levels of phosphorylated serine 65 of ubiquitin and PRKN have been observed, which are associated with the phosphorylation of ubiquitin by PINK1 at the outer mitochondrial membrane (OMM), have been identified in postmortem PD brains (107). These investigations substantiate a notable association between neuroinflammation and the activation of the PINK1/Parkin pathway in PD, indicating that mitochondrial autophagy has a pivotal role in averting neuroinflammation within this pathological framework.
The involvement of α-synuclein in perturbing mitochondrial function has been previously substantiated (108). It has been documented that α-synuclein possesses a mitochondrial targeting sequence at its N-terminal region, allowing its localization to the OMM. This localization facilitates interactions with components of the outer membrane receptors, thereby leading to compromised cellular respiration (109). This phenomenon has been observed in models of PD and in post-mortem brain tissue from individuals with PD (108,110). Certain α-synuclein species have the capability to intricately bind with the translocase of outer mitochondrial membrane 20 receptor, contributing to mitochondrial dysfunction and an elevated generation of ROS (108). Notably, a study employed a seeding-based model of α-synuclein fibrillization to validate that the progression of LB formation, beyond mere fibril assembly, is a key catalyst in neurodegeneration, additionally exacerbating mitochondrial impairment and synaptic dysfunction (111). This suggests an intrinsic link between mitochondrial dysfunction, α-synuclein aggregation and the formation of LBs.
LRRK2 is another pivotal gene implicated in mitochondrial dysfunction within the context of PD. This gene exerts its influence by modulating the OMM adaptor protein responsible for orchestrating mitophagy, a critical process in maintaining mitochondrial quality. Consequently, this regulatory role of LRRK2 leads to diminished mitochondrial transport along the intricate cytoskeletal network (112). Consistent findings were observed in neurons from LRRK2 mutant rats (113). Mutant LRRK2 inhibits the recruitment of Parkin to the OMM and its interaction with Drp1, thereby suppressing PINK1/Parkin-mediated autophagy, which leads to impaired segregation and degradation of damaged mitochondria (114).
The NLRP3 inflammasome assumes a pivotal role in instigating the neuroinflammatory cascade observed in PD. Heightened levels of inflammasome constituents and inflammation-associated factors have been discerned within blood samples sourced from individuals with PD (105,115). Mitochondrial impairment within microglia, coupled with the activation of the NLRP3 inflammasome, has been reported in both in vitro and in vivo models of PD (116). In addition, activation of NLRP3 has been observed in PINK1−/− or Parkin−/− microglia, while inhibitors of the inflammasome can effectively suppress this activation process (117). Furthermore, the attenuation of NLRP3 inflammasome activation not only mitigates neuroinflammation and ameliorates motor impairments but also safeguards against the depletion of DA neurons in both a mPTP-induced PD model and a human α-synuclein overexpression PD model (118).
Collectively, the convergence of α-synuclein oligomerization, genetic mutations, impaired mitochondrial autophagy and NLRP3 activation constitutes a synergistic interplay contributing to mitochondria-associated neuroinflammation during the progression of PD (Fig. 3).
MS
MS is a chronic inflammatory disease of the CNS characterized by demyelination and axonal degeneration (119). The inflammation observed in MS arises from elements of both the innate and adaptive immune systems, encompassing the proliferation and dysregulation of pro-inflammatory T lymphocytes, activation of B cells and secretion of inflammatory cytokines (120). At the onset of MS, pathogenic inflammatory T lymphocytes infiltrate the CNS, triggering an immune response that activates microglia and astrocytes, leading to acute inflammation. Subsequently, B cells are further activated, initiating a cascade that sustains chronic inflammation (121). While anti-inflammatory and immunomodulatory therapies have become mainstream in the treatment of acute demyelinating episodes, options remain limited for addressing the progressive stages of MS (122). Further exploration of the pathogenesis of MS is therefore still required.
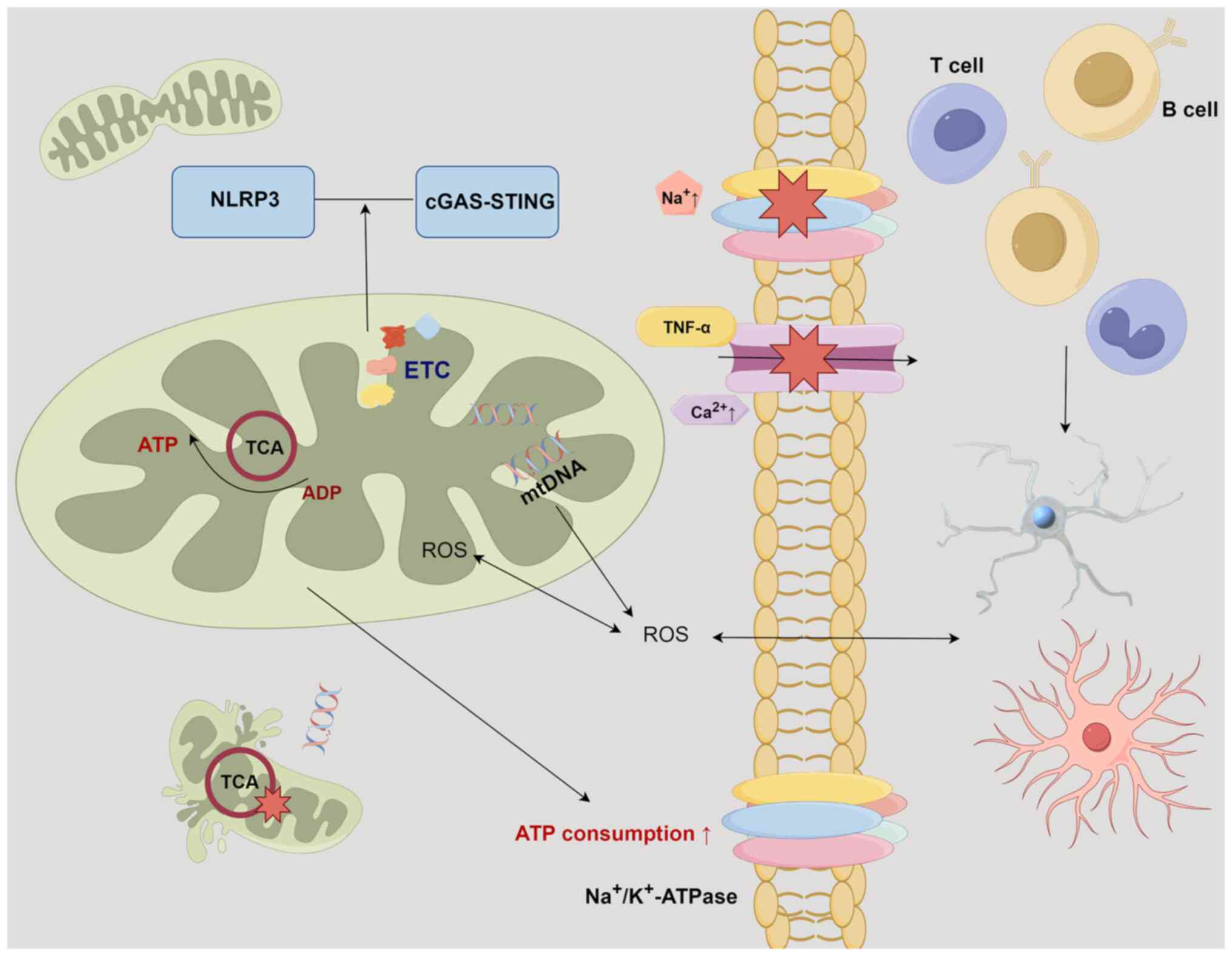
A recent study has substantiated that mitochondrial dysfunction contributes to CNS damage in MS (123). Mitochondria function as the principal energy supply units within neurons. Neurons facilitate signal transmission through membrane depolarization, which is facilitated by the electrochemical gradient of Na+/K+-ATPase. In the context of MS, the interplay of chronic inflammation and myelin disruption leads to a redistribution of ion channels. The heightened presence of Na+/K+-ATPase intensifies ATP consumption (123). At this critical juncture, mitochondria can compensate by augmenting both their quantity and volume, thereby inducing alterations in neuron positioning and morphology (123). Persistent inflammation triggers the activation of macrophages and microglial cells, thereby instigating the release of ROS and inducing oxidative stress (124). This exacerbates the release of glutamate, ultimately culminating in neuronal damage (120). Oxidative stress imposes secondary damage on both mitochondria and macromolecules (such as mtDNA, ETC proteins and lipids), thereby significantly impairing energy generation (120). While nuclear factor erythroid 2-related factor 2 and antioxidant enzymes such as heme oxygenase-1, are activated during periods of hypoxic stress to compensate for mitochondrial dysfunction, once a critical threshold of reduced ATP production is reached, ion homeostasis becomes compromised (125). This disruption results in chronic inflammation and triggers Ca2+-dependent proteases, ultimately leading to apoptosis within demyelinated axons (126). The presence of oxidized DNA and lipids has been observed in apoptotic oligodendrocytes and dystrophic axons within active MS lesions (127). Furthermore, the inflammatory factor, TNF-α, exacerbates the impairment of OXPHOS through Ca2+ modulation (128). The resultant reduction in ATP production hampers the ability of the Na+/K+-ATPase to maintain gradients after action potentials, leading to an accumulation of sodium within the neuronal cytoplasm. This phenomenon, in turn, compels Na+/Ca2+channels to facilitate intracellular calcium transfer, initiating a cascade of Ca2+-dependent apoptosis that ultimately culminates in neuronal death. This intricate process significantly contributes to Wallerian degeneration and irreversible neurofunctional impairment (120). In MS animal models, double-strand breaks in mtDNA lead to chronic demyelination and axonal degeneration, which are exacerbated over time (129). It is noteworthy that mitochondrial dysfunction and oligodendrocyte myelin formation are inherently interconnected. The level of the mitochondrial metabolite, N-acetylaspartate (NAA), is reduced in the normal-appearing white matter of patients with MS (130,131). In vitro experiments have confirmed that extracellular NAA improves Oli-neuM cell differentiation and axonal connectivity (132). Furthermore, the NLRP3 inflammasome and cyclic GMP-AMP synthase-STING pathway, which are associated with increased mitochondrial damage and respiratory stress, are activated in MS (133,134). These findings highlight the pivotal role of mitochondrial function in the progression of MS (Fig. 4).
Epilepsy
Epilepsy is a persistent neurological condition distinguished by the recurrence of seizures intertwined with an underlying neurodegenerative process (135). The intricate diversity of epilepsy presents a significant challenge for its treatment (136). Moreover, the success rate of antiepileptic drug therapy remains limited, ranging from 30 to 50% (137). Lately, there has been growing interest in the role of oxidative stress and redox dysregulation in epilepsy. Elevated levels of diverse biomarkers associated with oxidative stress and neuroinflammation have been reported in the brains and peripheral tissues of both human patients and animal epilepsy models (138,139). Therefore, anti-inflammatory and antioxidant therapies hold promising therapeutic potential. Administering these treatments shortly before or after the symptomatic onset of epilepsy could effectively hinder the advancement of spontaneous seizures and potentially delay their onset (140). Moreover, it has been demonstrated that IL-4 exerts a neuroprotective effect during epileptogenesis by lowering TNF-α levels and mitigating mitochondrial swelling in a mouse model induced by kaliotoxin (141).
Mitochondrial dysfunction has a pivotal role in the connection between epilepsy and oxidative stress (136). In total, ~40% of individuals with epilepsy exhibit concomitant mitochondrial disorders (142). Mitochondrial dysfunction disrupts the balance of RNS and ROS, resulting in heightened ROS generation, oxidative harm and diminished ATP production. This cascade ultimately culminates in mtDNA mutations and compromised mitochondrial respiration, establishing a detrimental cycle (1,136). Furthermore, there was a notable rise in the occurrence of spontaneous motor seizures within mitochondrial SOD2−/− mice (143). Conditional deletion of SOD2 specifically in the forebrain led to reduced mitochondrial oxygen consumption and the subsequent development of epilepsy in mice (42). An additional study demonstrated that the targeted removal of neuron-specific mitochondrial SOD2 results in a severe and intricate epileptic phenotype (144). Collectively, these findings indicate that mitochondrial oxidative stress is not solely a result of epilepsy but also contributes to its onset.
Additionally, ROS can directly regulate pro-inflammatory molecules, including IL-1β, high mobility group box-1 (HMGB1) and matrix metalloproteinase 9 (136). Pathways related to HMGB1, toll-like receptor 4 (TLR4) and IL-1β/interleukin-1 receptor 1 have therefore emerged as potential targets for epilepsy therapy (145). HMGB1 interacts with TLR4 and functions as a proinflammatory cytokine in the extracellular environment. Research has demonstrated that the translocation of nuclear HMGB1 and active caspase-3 to the mitochondria enhances programmed necrotic cell death in parvalbumin cells and CA1 neurons during status epilepticus (146-148). Furthermore, another study discovered that the levels of glutathione, an essential antioxidant for maintaining mitochondrial integrity, significantly increased following the administration of antioxidant drugs [N-acetylcysteine (NAC) and sulforaphane]. Concurrently, this intervention led to a reduction in HMGB1 production within an acquired epilepsy rat model induced by status epilepticus (149,150). The NLRP3 inflammasome is a molecule associated with epilepsy, which can be triggered by ROS (151,152). Furthermore, it has been empirically shown that mtROS function as a secondary messenger, triggering the activation of NLRP3 and its translocation to the mitochondria, thereby facilitating the activation of IL-1β. Consequently, this process elicits a proinflammatory signal in reaction to mitochondrial dysfunction (153,154). The available evidence suggests that activation of the NLRP3 inflammasome can be inhibited by anti-inflammatory and antioxidant therapy, thereby potentially influencing epileptogenesis (155). A previous study also observed upregulated levels of NLRP3 and IL-1β in children diagnosed with febrile seizures (156). It has also been suggested that NLRP1 and NLRP4 may have roles in the development of epilepsy. Specifically, NLRP1 has been found to be upregulated in patients with temporal lobe epilepsy (TLE) (157). In a TLE rat model, reducing the expression of NLRP1 was shown to decrease the frequency and severity of seizures (158). The sensitivity of domain-containing protein 4 (NLRC4) to mtROS in astrocytes and its association with mitochondrial oxidative stress in neurodegenerative diseases provide insights into the potential mechanism of NLRC4 in epilepsy (159). This highlights the intricate relationship between epilepsy and mitochondrial dysfunction, emphasizing the need to unravel the multifaceted impact of mitochondrial function on brain activity as a potential avenue for epilepsy treatment.
4. Therapies targeting mitochondria
Mitochondrial therapy for neuroinflammation is an emerging field with potential for the treatment of chronic neuroinflammatory diseases (160). Currently, the overall treatment strategy includes restoring normal mitochondrial physiological functions (ATP production) and antioxidant therapy, clearing mitochondria with abnormal functions through mitochondrial autophagy or other mitochondrial stress responses, gene therapy, restoring mitochondrial dynamics and addressing mitochondrial calcium ion balance disorders (55,149,160-162).
Cytokines such as IL-1-β and TNF-α, as well as their receptors, serve a significant role in the development and progression of AD (55). Inhibiting the interaction of these pro-inflammatory factors may therefore be a more effective therapeutic option for AD. For instance, the anti-inflammatory molecule, minocycline, reduces Aβ and τ pathological lesions in an AD rat model by inhibiting pro-inflammatory cytokines in glial cells through the NF-κB signaling pathway (55). Inhibiting inducible NO synthase and cyclooxygenase-2 is also considered to effectively improve neuroinflammation in patients with AD (163). Excessive activation of microglial cells and reduced phagocytic ability leads to increased accumulation of Aβ plaques and τ hyperphosphorylation, exacerbating neuroinflammation (164). Microglial polarization from an M1 state to a neuroprotective M2 state is also a potential target for treatment. GV-971, a sodium oligomannate, modulates gut microbiota amino acid metabolism to reduce the activation of T helper 1 cells, thereby inhibiting M1-type microglia activation, ultimately alleviating neuroinflammation and enhancing cognitive function in AD mice (165). The second-generation tetracycline, minocycline, selectively inhibits the M1 state of microglial cells and exerts anti-neuroinflammatory effects in patients with AD (166). Antioxidants are also extensively researched in drug development. For instance, vitamin E reduces the production of TNF-α and NO, lowering the levels of ROS and IL-6 induced by lipopolysaccharides in microglial cells, thereby providing neuroprotection (167). Polyphenolic compounds such as flavonoids and vitamin C, may help prevent age-related neurodegenerative diseases based on a clinical study (168). Flavonoids, which are found in daily dietary products, promote the survival of neurons in patients with AD by reducing protein oxidation, inhibiting the JNK and p38 pathways and preventing the production of free radicals (168).
Mitochondrial dysfunction is a well-established feature of PD, with defects in mitochondrial complex I activity and increased oxidative stress (169). In response to the characteristics of this disease, the application of the mitochondria-targeted antioxidant, mitoquinone (MitoQ), has been gradually gaining attention. Cell experiments have confirmed that MitoQ can reduce membrane leakage, oxidative stress and apoptosis induced by α-synuclein (170). In fruit flies with PINK1 knockout, vitamin K2, structurally similar to coenzyme Q10 and also serving as an electron carrier in the ETC, was found to alleviate oxidative stress in PD (162). However, in a double-blind clinical study assessing untreated patients with PD using the Unified Parkinson Disease Rating Scale, it was discovered that PD did not improve after 12 months of MitoQ administration (171). Further research is therefore needed to determine the effectiveness of MitoQ. Niacinamide (Vitamin B3 and NAM) and its derivatives are currently under investigation, with the aim to normalize redox levels (172). NAC has also been reported to have demonstrated antioxidant properties in a clinical trial (173). Additionally, a promising candidate in recent clinical trials is ursodeoxycholic acid (UDCA), known for its broad safety profile and its ability to prevent mitochondrial membrane depolarization and stabilize cytochrome c in mitochondria (174,175). The therapeutic potential of UDCA in treating mitochondrial damage has been demonstrated in LRRK2G2019S mutant PD patients and LRRK2G2019S transgenic flies (161). Mitochondrial autophagy is also a crucial target for PD treatment. In PD cell and mouse models, celastrol plays a neuroprotective role by activating mitochondrial autophagy and inhibiting DA neuron loss (176). Furthermore, mitochondrial dynamics may represent a potential target for PD treatment. A study has confirmed that the mitochondrial fission GTPase Drp1 inhibitor, mdivi-1, can be used to inhibit mitochondrial fragmentation in α-synuclein rat PD models, reducing neurodegeneration and mitochondrial oxidative stress (177). Notably, it has been observed in both animal models and patients with PD that physical exercise can enhance mitochondrial biogenesis, providing new avenues for the treatment of PD (178,179).
The treatment of MS is inherently complex due to the varying subtypes of MS, each requiring distinct therapeutic approaches. While significant progress has been made in the treatment of MS, such as the effectiveness of the anti-CD20 antibody, ocrelizumab, and the sphingosine-1-phosphate receptor (S1PR) modulator, siponimod, in patients with primary progressive MS and relapsing-remitting MS (180,181), the management of other progressive forms of MS remains challenging. For instance, in phase III clinical trials, the S1PR modulator, fingolimod, did not demonstrate a reduction in disability progression in patients with primary progressive MS (182). Immune-modulating compounds, such as siponimod and ocrelizumab, targeting degenerative mechanisms may therefore not comprehensively address neurodegenerative processes. Furthermore, the development of new drugs is hindered by the incomplete understanding of the pathogenesis of progressive MS and the absence of suitable animal models. The onset of MS is often associated with the activation of microglia and the continued involvement of T cells and B cells, which release high levels of ROS and RNS, leading to mitochondrial and axonal damage, and ultimately resulting in neurodegeneration. Therefore, targeting mitochondria has emerged as a focal point in the elucidation of methods to combat MS. Currently, mitochondrial protective strategies, such as minocycline, iron (Fe2+) chelating compounds and antioxidants that reduce oxidative stress, have shown a certain degree of efficacy in MS treatment (119). Recent research has elucidated that mitochondrial dysfunction impairs Na+/K+-ATPase, leading to Na+/Ca2+ exchanger reversal and calcium overload, thereby mediating axonal degeneration (183). Notably, mitochondrial transplantation into the medial forebrain bundle has been shown to ameliorate motor deficits in 6-hydroxydopamine-induced PD rats, enhancing mitochondrial functionality (184). It has also been demonstrated that neural stem cells effectively deliver functional mitochondria to target cells via extracellular vesicles, thereby remedying mitochondrial functional deficits in mice with experimental autoimmune encephalomyelitis (184). These studies therefore provide evidence supporting the potential use of mitochondrial transplantation as a therapeutic strategy for MS in the future.
Mitochondrial dysfunction is one of the most prominent features of epilepsy, affecting 35-60% of patients with epilepsy (185). Previous research has found that cannabidiol (CBD) can reduce the frequency of epileptic seizures (186). This may be related to its ability to induce the formation of mitochondrial-derived vesicles through the PINK1/Parkin pathway, which participates in mitochondrial repair (187). Recent evidence suggests that CBD engages in mitochondrial-related anti-inflammatory and antioxidant activities, where it reverses iron-induced mitochondrial dysfunction by rescuing mitochondrial ferritin and modulating mtDNA epigenetics, and participates in neurodegenerative mechanisms via the NF-κB, phosphorylated p38 MAPK and peroxisome proliferation-activated receptor γ pathways (188). Currently, the Food and Drug Administration has approved the drug compound, Epidiolex, which contains CBD, for the treatment of seizures (189). Furthermore, a study has also found that the IL-1 receptor antagonist, anakinra, can reduce seizure frequency (190). The antiepileptic drug, levetiracetam, can reduce neuronal excitability by restoring the resting membrane potentials of IL-1β-induced neurotoxic astrocytes and promoting the secretion of TGF-β1 (191). Additionally, levetiracetam modulates the opening of the mPTP via synaptic vesicle protein 2A, reducing neural hyperexcitability in patients with AD and AD animal models (192). Other antioxidants targeting mitochondria, such as polyphenols, vitamins and thiols, have been shown to help reduce epileptic seizures (193). Therefore, targeting mitochondria may be a key approach to treating epilepsy.
In general, the treatment strategies for chronic neuroinflammatory diseases remain focused on combating oxidative stress and ameliorating mitochondrial functional impairments, which constitute shared pathophysiological features of such conditions. Mitochondrially-targeted therapy stands out among emerging therapeutic modalities due to its ability to selectively target mitochondria, neutralizing reactive ROS and restoring their functionality. Currently, research on agents such as MitoQ is the most extensive. However, despite demonstrating significant therapeutic effects in animal models, MitoQ has not yielded the anticipated substantial benefits in clinical trials for PD or AD (194,195). Moreover, effectively penetrating the blood-brain barrier and achieving optimal concentrations within target brain tissues remain pressing challenges. Additionally, mitochondrial-targeted therapies fail to selectively recognize damaged mitochondria and cannot directly modulate mitochondrial dynamics and mitophagy processes, which may contribute to their suboptimal clinical efficacy (194). In addition to mitochondrial-targeted therapy, mitochondrial gene therapy is emerging as a novel research domain. Treatment strategies encompass restoring normal mitochondrial function, repairing or eliminating mutated mtDNA and delivering wild-type mtDNA (196). Despite the development of various delivery systems, including mitochondria-targeting peptides and liposomes, as well as physical methods such as electroporation and hydrodynamic injection, effectively delivering therapeutic macromolecules to mitochondria remains a challenge due to the presence of the blood-brain barrier (196). Furthermore, delivery systems may induce cytotoxicity or interact with endogenous biomolecules, leading to aggregation and reduced efficacy (197). The development of mitochondrial genome editing technology is still in its nascent stages, necessitating further understanding of how RNA and editing tools penetrate mammalian mitochondria (198). Efforts to develop more precise and safer mitochondrial-targeted drugs may therefore be a future research focus.
5. Conclusions and future directions
Mitochondrial dysfunction is a common feature of chronic neuroinflammatory diseases and exploring its pathological mechanisms may provide new avenues for future treatments. Various drugs have been developed that target mitochondria, focusing on aspects such as antioxidation, mitochondrial autophagy regulation, calcium ion balance and gene repair. However, clinical application of these drugs remains a significant challenge. Exploring new therapeutic targets, selectively targeting dysfunctional mitochondria, ensuring delivery of drugs across the blood-brain barrier into the brain and minimizing adverse reactions may be the focus of future research. Additionally, advancements in mitochondrial genome editing technology offer hope for the precise manipulation of mitochondrial function and addressing genetic abnormalities in neuroinflammatory diseases.
Future treatment strategies may not be limited to a single approach; combining anti-inflammatory and antioxidative therapy with mitochondrial-targeted treatment may enhance overall treatment safety and efficacy. In conclusion, while mitochondrial-targeted therapy holds promise for the treatment of chronic neuroinflammatory diseases, addressing current limitations is crucial. By overcoming delivery challenges, enhancing treatment specificity and exploring new therapeutic targets, mitochondrial-targeted therapy holds promise for treating chronic neuroinflammatory diseases and other neurological disorders.
Availability of data and materials
Not applicable.
Authors' contributions
PQ and YS contributed equally to this work. Conceptualization, writing the original draft and reviewing and editing the manuscript was conducted by LL; writing the original draft and reviewing the manuscript was conducted by PQ; Reviewing and editing the manuscript and drawing the figures was conducted by YS. All authors read and approved the final version of the manuscript. Data authentication is not applicable.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Acknowledgments
Not applicable.
Funding
No funding was received.
References
|
Vezzani B, Carinci M, Patergnani S, Pasquin MP, Guarino A, Aziz N, Pinton P, Simonato M and Giorgi C: The Dichotomous role of inflammation in the CNS: A mitochondrial point of view. Biomolecules. 10:14372020. View Article : Google Scholar : PubMed/NCBI | |
|
Yilmaz C, Karali K, Fodelianaki G, Gravanis A, Chavakis T, Charalampopoulos I and Alexaki VI: Neurosteroids as regulators of neuroinflammation. Front Neuroendocrinol. 55:1007882019. View Article : Google Scholar : PubMed/NCBI | |
|
Fontana L, Ghezzi L, Cross AH and Piccio L: Effects of dietary restriction on neuroinflammation in neurodegenerative diseases. J Exp Med. 218:e201900862021. View Article : Google Scholar : PubMed/NCBI | |
|
Biswas K: Microglia mediated neuroinflammation in neurodegenerative diseases: A review on the cell signaling pathways involved in microglial activation. J Neuroimmunol. 383:5781802023. View Article : Google Scholar : PubMed/NCBI | |
|
Liu A, Yu L, Li X, Zhang K, Zhang W, So KF, Tissir F, Qu Y and Zhou L: Celsr2-mediated morphological polarization and functional phenotype of reactive astrocytes in neural repair. Glia. 71:1985–2004. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Fan YY and Huo J: A1/A2 astrocytes in central nervous system injuries and diseases: Angels or devils? Neurochem Int. 148:1050802021. View Article : Google Scholar : PubMed/NCBI | |
|
Singh D: Astrocytic and microglial cells as the modulators of neuroinflammation in Alzheimer's disease. J Neuroinflammation. 19:2062022. View Article : Google Scholar : PubMed/NCBI | |
|
Gimenez MA, Sim J, Archambault AS, Klein RS and Russell JH: A tumor necrosis factor receptor 1-dependent conversation between central nervous system-specific T cells and the central nervous system is required for inflammatory infiltration of the spinal cord. Am J Pathol. 168:1200–1209. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Da Mesquita S, Fu Z and Kipnis J: The meningeal lymphatic system: A new player in neurophysiology. Neuron. 100:375–388. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Flores-Romero H, Dadsena S and Garcia-Saez AJ: Mitochondrial pores at the crossroad between cell death and inflammatory signaling. Mol Cell. 83:843–856. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Wei Y, Miao Q, Zhang Q, Mao S, Li M, Xu X, Xia X, Wei K, Fan Y, Zheng X, et al: Aerobic glycolysis is the predominant means of glucose metabolism in neuronal somata, which protects against oxidative damage. Nat Neurosci. 26:2081–2089. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Rose J, Brian C, Woods J, Pappa A, Panayiotidis MI, Powers R and Franco R: Mitochondrial dysfunction in glial cells: Implications for neuronal homeostasis and survival. Toxicology. 391:109–115. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Rizzuto R, De Stefani D, Raffaello A and Mammucari C: Mitochondria as sensors and regulators of calcium signalling. Nat Rev Mol Cell Biol. 13:566–578. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Satarker S, Bojja SL, Gurram PC, Mudgal J, Arora D and Nampoothiri M: Astrocytic glutamatergic transmission and its implications in neurodegenerative disorders. Cells. 11:11392022. View Article : Google Scholar : PubMed/NCBI | |
|
Morales-Ropero JM, Arroyo-Urea S, Neubrand VE, Martín-Oliva D, Marín-Teva JL, Cuadros MA, Vangheluwe P, Navascués J, Mata AM and Sepúlveda MR: The endoplasmic reticulum Ca(2+) -ATPase SERCA2b is upregulated in activated microglia and its inhibition causes opposite effects on migration and phagocytosis. Glia. 69:842–857. 2021. View Article : Google Scholar | |
|
Neel DV, Basu H, Gunner G, Bergstresser MD, Giadone RM, Chung H, Miao R, Chou V, Brody E, Jiang X, et al: Gasdermin-E mediates mitochondrial damage in axons and neurodegeneration. Neuron. 111:1222–1240 e1229. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Borsche M, Pereira SL, Klein C and Grünewald A: Mitochondria and Parkinson's disease: Clinical, molecular, and translational aspects. J Parkinsons Dis. 11:45–60. 2021. View Article : Google Scholar : | |
|
Hinkle JT, Patel J, Panicker N, Karuppagounder SS, Biswas D, Belingon B, Chen R, Brahmachari S, Pletnikova O, Troncoso JC, et al: STING mediates neurodegeneration and neuroinflammation in nigrostriatal α-synucleinopathy. Proc Natl Acad Sci USA. 119:e21188191192022. View Article : Google Scholar | |
|
Pezone A, Olivieri F, Napoli MV, Procopio A, Avvedimento EV and Gabrielli A: inflammation and DNA damage: Cause, effect or both. Nat Rev Rheumatol. 19:200–211. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Duarte JN: Neuroinflammatory mechanisms of mitochondrial dysfunction and neurodegeneration in glaucoma. J Ophthalmol. 2021:45819092021.PubMed/NCBI | |
|
Yang Y, Liu Y, Zhu J, Song S, Huang Y, Zhang W, Sun Y, Hao J, Yang X, Gao Q, et al: Neuroinflammation-mediated mitochondrial dysregulation involved in postoperative cognitive dysfunction. Free Radic Biol Med. 178:134–146. 2022. View Article : Google Scholar | |
|
Pan RY, Ma J, Kong XX, Wang XF, Li SS, Qi XL, Yan YH, Cheng J, Liu Q, Jin W, et al: Sodium rutin ameliorates Alzheimer's disease-like pathology by enhancing microglial amyloid-β clearance. Sci Adv. 5:eaau63282019. View Article : Google Scholar | |
|
Joshi AU, Minhas PS, Liddelow SA, Haileselassie B, Andreasson KI, Dorn GW II and Mochly-Rosen D: Fragmented mitochondria released from microglia trigger A1 astrocytic response and propagate inflammatory neurodegeneration. Nat Neurosci. 22:1635–1648. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Mi Y, Qi G, Vitali F, Shang Y, Raikes AC, Wang T, Jin Y, Brinton RD, Gu H and Yin F: Loss of fatty acid degradation by astrocytic mitochondria triggers neuroinflammation and neurodegeneration. Nat Metab. 5:445–465. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Cenini G, Rub C, Bruderek M and Voos W: Amyloid β-peptides interfere with mitochondrial preprotein import competence by a coaggregation process. Mol Biol Cell. 27:3257–3272. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Bingol B, Tea JS, Phu L, Reichelt M, Bakalarski CE, Song Q, Foreman O, Kirkpatrick DS and Sheng M: The mitochondrial deubiquitinase USP30 opposes parkin-mediated mitophagy. Nature. 510:370–375. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Annesley SJ and Fisher PR: Mitochondria in health and disease. Cells. 8:6802019. View Article : Google Scholar : PubMed/NCBI | |
|
Lin MT and Beal MF: Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature. 443:787–795. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Johnson J, Mercado-Ayon E, Mercado-Ayon Y, Dong YN, Halawani S, Ngaba L and Lynch DR: Mitochondrial dysfunction in the development and progression of neurodegenerative diseases. Arch Biochem Biophys. 702:1086982021. View Article : Google Scholar | |
|
Adebayo M, Singh S, Singh AP and Dasgupta S: Mitochondrial fusion and fission: The fine-tune balance for cellular homeostasis. FASEB J. 35:e216202021. View Article : Google Scholar : PubMed/NCBI | |
|
Alevriadou BR, Patel A, Noble M, Ghosh S, Gohil VM, Stathopulos PB and Madesh M: Molecular nature and physiological role of the mitochondrial calcium uniporter channel. Am J Physiol Cell Physiol. 320:C465–C482. 2021. View Article : Google Scholar : | |
|
Fischer R and Maier O: Interrelation of oxidative stress and inflammation in neurodegenerative disease: Role of TNF. Oxid Med Cell Longev. 2015:6108132015. View Article : Google Scholar : PubMed/NCBI | |
|
Teleanu DM, Niculescu AG, Lungu II, Radu CI, Vladâcenco O, Roza E, Costăchescu B, Grumezescu AM and Teleanu RI: An overview of oxidative stress, neuroinflammation, and neurodegenerative diseases. Int J Mol Sci. 23:59382022. View Article : Google Scholar : PubMed/NCBI | |
|
Tsang T, Davis CI and Brady DC: Copper biology. Curr Biol. 31:R421–R427. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Borisov VB, Siletsky SA, Nastasi MR and Forte E: ROS defense systems and terminal oxidases in bacteria. Antioxidants (Basel). 10:8392021. View Article : Google Scholar : PubMed/NCBI | |
|
Haider S, Batool Z, Ahmad S, Siddiqui RA and Haleem DJ: Walnut supplementation reverses the scopolamine-induced memory impairment by restoration of cholinergic function via mitigating oxidative stress in rats: A potential therapeutic intervention for age related neurodegenerative disorders. Metab Brain Dis. 33:39–51. 2018. View Article : Google Scholar | |
|
Luque-Contreras D, Carvajal K, Toral-Rios D, Franco-Bocanegra D and Campos-Pena V: Oxidative stress and metabolic syndrome: Cause or consequence of Alzheimer's disease? Oxid Med Cell Longev. 2014:4978022014. View Article : Google Scholar : PubMed/NCBI | |
|
Singh A, Kukreti R, Saso L and Kukreti S: Oxidative stress: A key modulator in neurodegenerative diseases. Molecules. 24:15832019. View Article : Google Scholar : PubMed/NCBI | |
|
Bhatia V and Sharma S: Role of mitochondrial dysfunction, oxidative stress and autophagy in progression of Alzheimer's disease. J Neurol Sci. 421:1172532021. View Article : Google Scholar : PubMed/NCBI | |
|
Zorov DB, Juhaszova M and Sollott SJ: Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol Rev. 94:909–950. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Farkhondeh T, Mehrpour O, Forouzanfar F, Roshanravan B and Samarghandian S: Oxidative stress and mitochondrial dysfunction in organophosphate pesticide-induced neurotoxicity and its amelioration: A review. Environ Sci Pollut Res Int. 27:24799–24814. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Rowley S, Liang LP, Fulton R, Shimizu T, Day B and Patel M: Mitochondrial respiration deficits driven by reactive oxygen species in experimental temporal lobe epilepsy. Neurobiol Dis. 75:151–158. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Bordoni L and Gabbianelli R: Mitochondrial DNA and neurodegeneration: Any role for dietary antioxidants? Antioxidants (Basel). 9:7642020. View Article : Google Scholar : PubMed/NCBI | |
|
Coppede F: Mitochondrial DNA methylation and mitochondria-related epigenetics in neurodegeneration. Neural Regen Res. 19:405–406. 2024. View Article : Google Scholar | |
|
Sharma VK, Singh TG and Mehta V: Stressed mitochondria: A target to intrude alzheimer's disease. Mitochondrion. 59:48–57. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Lin J and Epel E: Stress and telomere shortening: Insights from cellular mechanisms. Ageing Res Rev. 73:1015072022. View Article : Google Scholar : | |
|
Ortiz JM and Swerdlow RH: Mitochondrial dysfunction in Alzheimer's disease: Role in pathogenesis and novel therapeutic opportunities. Br J Pharmacol. 176:3489–3507. 2019. View Article : Google Scholar | |
|
Liu H, Zhang H, Zhang Y, Xu S, Zhao H, He H and Liu X: Modeling mtDNA hypermethylation vicious circle mediating Aβ-induced endothelial damage memory in HCMEC/D3 cell. Aging (Albany NY). 12:18343–18362. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Rasheed M, Liang J, Wang C, Deng Y and Chen Z: Epigenetic regulation of neuroinflammation in Parkinson's disease. Int J Mol Sci. 22:49562021. View Article : Google Scholar : PubMed/NCBI | |
|
Youle RJ and van der Bliek AM: Mitochondrial fission, fusion, and stress. Science. 337:1062–1065. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Song Y, Xu Y, Liu Y, Gao J, Feng L, Zhang Y, Shi L and Zhang M, Guo D, Qi B and Zhang M: Mitochondrial quality control in the maintenance of cardiovascular homeostasis: The roles and interregulation of UPS, mitochondrial dynamics and mitophagy. Oxid Med Cell Longev. 2021:39607732021. View Article : Google Scholar : PubMed/NCBI | |
|
Xie JH, Li YY and Jin J: The essential functions of mitochondrial dynamics in immune cells. Cell Mol Immunol. 17:712–721. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Chakrabarti S and Bisaglia M: Oxidative stress and neuroinflammation in Parkinson's disease: The role of dopamine oxidation products. Antioxidants (Basel). 12:9552023. View Article : Google Scholar : PubMed/NCBI | |
|
Picca A, Ferri E, Calvani R, Coelho-Junior HJ, Marzetti E and Arosio B: Age-Associated glia remodeling and mitochondrial dysfunction in neurodegeneration: Antioxidant supplementation as a possible intervention. Nutrients. 14:24062022. View Article : Google Scholar : PubMed/NCBI | |
|
Dhapola R, Hota SS, Sarma P, Bhattacharyya A, Medhi B and Reddy DH: Recent advances in molecular pathways and therapeutic implications targeting neuroinflammation for Alzheimer's disease. Inflammopharmacology. 29:1669–1681. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Chang YH, Lin HY, Shen FC, Su YJ, Chuang JH, Lin TK, Liou CW, Lin CY, Weng SW and Wang PW: The causal role of mitochondrial dynamics in regulating innate immunity in diabetes. Front Endocrinol (Lausanne). 11:4452020. View Article : Google Scholar : PubMed/NCBI | |
|
Paik S, Kim JK, Silwal P, Sasakawa C and Jo EK: An update on the regulatory mechanisms of NLRP3 inflammasome activation. Cell Mol Immunol. 18:1141–1160. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Yurtsever I, Ustundag UV, Ünal I, Ateş PS and Emekli-Alturfan E: Rifampicin decreases neuroinflammation to maintain mitochondrial function and calcium homeostasis in rotenone-treated zebrafish. Drug Chem Toxicol. 45:1544–1551. 2022. View Article : Google Scholar | |
|
Kannurpatti SS: Mitochondrial calcium homeostasis: Implications for neurovascular and neurometabolic coupling. J Cereb Blood Flow Metab. 37:381–395. 2017. View Article : Google Scholar : | |
|
Poewe W, Seppi K, Tanner CM, Halliday GM, Brundin P, Volkmann J, Schrag AE and Lang AE: Parkinson disease. Nat Rev Dis Primers. 3:170132017. View Article : Google Scholar : PubMed/NCBI | |
|
Binvignat O and Olloquequi J: Excitotoxicity as a target against neurodegenerative processes. Curr Pharm Des. 26:1251–1262. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Yao L, Wu J, Koc S and Lu G: Genetic imaging of neuroinflammation in Parkinson's disease: Recent advancements. Front Cell Dev Biol. 9:6558192021. View Article : Google Scholar : PubMed/NCBI | |
|
Obrador E, Salvador R, Lopez-Blanch R, Jihad-Jebbar A, Valles SL and Estrela JM: Oxidative stress, neuroinflammation and mitochondria in the pathophysiology of amyotrophic lateral sclerosis. Antioxidants (Basel). 9:9012020. View Article : Google Scholar : PubMed/NCBI | |
|
Cyrino LAR, Delwing-de Lima D, Ullmann OM and Maia TP: Concepts of neuroinflammation and their relationship with impaired mitochondrial functions in bipolar disorder. Front Behav Neurosci. 15:6094872021. View Article : Google Scholar : PubMed/NCBI | |
|
Verkhratsky A, Rodriguez-Arellano JJ, Parpura V and Zorec R: Astroglial calcium signalling in Alzheimer's disease. Biochem Biophys Res Commun. 483:1005–1012. 2017. View Article : Google Scholar | |
|
Casaril AM, Katsalifis A, Schmidt RM and Bas-Orth C: Activated glia cells cause bioenergetic impairment of neurons that can be rescued by knock-down of the mitochondrial calcium uniporter. Biochem Biophys Res Commun. 608:45–51. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Garbincius JF and Elrod JW: Mitochondrial calcium exchange in physiology and disease. Physiol Rev. 102:893–992. 2022. View Article : Google Scholar : | |
|
Baumgartner HK, Gerasimenko JV, Thorne C, Ferdek P, Pozzan T, Tepikin AV, Petersen OH, Sutton R, Watson AJ and Gerasimenko OV: Calcium elevation in mitochondria is the main Ca2+ requirement for mitochondrial permeability transition pore (mPTP) opening. J Biol Chem. 284:20796–20803. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Green DR and Kroemer G: The pathophysiology of mitochondrial cell death. Science. 305:626–629. 2004. View Article : Google Scholar : PubMed/NCBI | |
|
Rimessi A, Previati M, Nigro F, Wieckowski MR and Pinton P: Mitochondrial reactive oxygen species and inflammation: Molecular mechanisms, diseases and promising therapies. Int J Biochem Cell Biol. 81:281–293. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Marchi S, Patergnani S, Missiroli S, Morciano G, Rimessi A, Wieckowski MR, Giorgi C and Pinton P: Mitochondrial and endoplasmic reticulum calcium homeostasis and cell death. Cell Calcium. 69:62–72. 2018. View Article : Google Scholar | |
|
Ooi K, Hu L, Feng Y, Han C, Ren X, Qian X, Huang H, Chen S, Shi Q, Lin H, et al: Sigma-1 receptor activation suppresses microglia M1 polarization via regulating endoplasmic reticulum-mitochondria contact and mitochondrial functions in stress-induced hypertension rats. Mol Neurobiol. 58:6625–6646. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Harland M, Torres S, Liu J and Wang X: Neuronal mitochondria modulation of LPS-induced neuroinflammation. J Neurosci. 40:1756–1765. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Lim D, Dematteis G, Tapella L, Genazzani AA, Calì T, Brini M and Verkhratsky A: Ca(2+) handling at the mitochondria-ER contact sites in neurodegeneration. Cell Calcium. 98:1024532021. View Article : Google Scholar : PubMed/NCBI | |
|
Krols M, van Isterdael G, Asselbergh B, Kremer A, Lippens S, Timmerman V and Janssens S: Mitochondria-associated membranes as hubs for neurodegeneration. Acta Neuropathol. 131:505–523. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Sunanda T, Ray B, Mahalakshmi AM, Bhat A, Rashan L, Rungratanawanich W, Song BJ, Essa MM, Sakharkar MK and Chidambaram SB: Mitochondria-endoplasmic reticulum crosstalk in Parkinson's disease: The role of brain renin angiotensin system components. Biomolecules. 11:16692021. View Article : Google Scholar : PubMed/NCBI | |
|
Du H, Guo L, Fang F, Chen D, Sosunov AA, McKhann GM, Yan Y, Wang C, Zhang H, Molkentin JD, et al: Cyclophilin D deficiency attenuates mitochondrial and neuronal perturbation and ameliorates learning and memory in Alzheimer's disease. Nat Med. 14:1097–1105. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Ayabe T, Takahashi C, Ohya R and Ano Y: β-Lactolin improves mitochondrial function in Abeta-treated mouse hippocampal neuronal cell line and a human iPSC-derived neuronal cell model of Alzheimer's disease. FASEB J. 36:e222772022. View Article : Google Scholar | |
|
Reddy PH and Beal MF: Amyloid beta, mitochondrial dysfunction and synaptic damage: Implications for cognitive decline in aging and Alzheimer's disease. Trends Mol Med. 14:45–53. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Dursun E, Alaylioglu M, Bilgic B, Hanağası H, Gürvit H, Emre M and Gezen-Ak D: Amyloid beta adsorption problem with transfer plates in amyloid beta 1-42 IVD Kits. J Mol Neurosci. 67:534–539. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Han J, Park H, Maharana C, Gwon AR, Park J, Baek SH, Bae HG, Cho Y, Kim HK, Sul JH, et al: Alzheimer's disease-causing presenilin-1 mutations have deleterious effects on mitochondrial function. Theranostics. 11:8855–8873. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Bai R, Guo J, Ye XY, Xie Y and Xie T: Oxidative stress: The core pathogenesis and mechanism of Alzheimer's disease. Ageing Res Rev. 77:1016192022. View Article : Google Scholar : PubMed/NCBI | |
|
Kowalczyk P, Sulejczak D, Kleczkowska P, Bukowska-Ośko I, Kucia M, Popiel M, Wietrak E, Kramkowski K, Wrzosek K and Kaczyńska K: Mitochondrial oxidative stress-A causative factor and therapeutic target in many diseases. Int J Mol Sci. 22:133842021. View Article : Google Scholar : PubMed/NCBI | |
|
Park MW, Cha HW, Kim J, Kim JH, Yang H, Yoon S, Boonpraman N, Yi SS, Yoo ID and Moon JS: NOX4 promotes ferroptosis of astrocytes by oxidative stress-induced lipid peroxidation via the impairment of mitochondrial metabolism in Alzheimer's diseases. Redox Biol. 41:1019472021. View Article : Google Scholar : PubMed/NCBI | |
|
Islam MT: Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol Res. 39:73–82. 2017. View Article : Google Scholar | |
|
Simpson DSA and Oliver PL: ROS generation in microglia: Understanding oxidative stress and inflammation in neurodegenerative disease. Antioxidants (Basel). 9:7432020. View Article : Google Scholar : PubMed/NCBI | |
|
ElAli A, Bordeleau M, Theriault P, Filali M, Lampron A and Rivest S: Tissue-plasminogen activator attenuates Alzheimer's disease-related pathology development in APPswe/PS1 mice. Neuropsychopharmacology. 41:1297–1307. 2016. View Article : Google Scholar : | |
|
Li Y, Xia X, Wang Y and Zheng JC: Mitochondrial dysfunction in microglia: A novel perspective for pathogenesis of Alzheimer's disease. J Neuroinflammation. 19:2482022. View Article : Google Scholar : PubMed/NCBI | |
|
Massey N, Shrestha D, Bhat SM, Kondru N, Charli A, Karriker LA, Kanthasamy AG and Charavaryamath C: Organic dust-induced mitochondrial dysfunction could be targeted via cGAS-STING or cytoplasmic NOX-2 inhibition using microglial cells and brain slice culture models. Cell Tissue Res. 384:465–486. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Ising C, Venegas C, Zhang S, Scheiblich H, Schmidt SV, Vieira-Saecker A, Schwartz S, Albasset S, McManus RM, Tejera D, et al: NLRP3 inflammasome activation drives tau pathology. Nature. 575:669–673. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Kelley N, Jeltema D, Duan Y and He Y: The NLRP3 inflammasome: An overview of mechanisms of activation and regulation. Int J Mol Sci. 20:33282019. View Article : Google Scholar : PubMed/NCBI | |
|
Jassim AH, Inman DM and Mitchell CH: Crosstalk between dysfunctional mitochondria and inflammation in glaucomatous neurodegeneration. Front Pharmacol. 12:6996232021. View Article : Google Scholar : PubMed/NCBI | |
|
Shaftel SS, Griffin WS and O'Banion MK: The role of interleukin-1 in neuroinflammation and Alzheimer disease: An evolving perspective. J Neuroinflammation. 5:72008. View Article : Google Scholar : PubMed/NCBI | |
|
Gonzalez-Reyes RE, Nava-Mesa MO, Vargas-Sanchez K, Ariza-Salamanca D and Mora-Munoz L: Involvement of astrocytes in Alzheimer's disease from a neuroinflammatory and oxidative stress perspective. Front Mol Neurosci. 10:4272017. View Article : Google Scholar | |
|
Sheng JG, Ito K, Skinner RD, Mrak RE, Rovnaghi CR, Van Eldik LJ and Griffin WS: In vivo and in vitro evidence supporting a role for the inflammatory cytokine interleukin-1 as a driving force in Alzheimer pathogenesis. Neurobiol Aging. 17:761–766. 1996. View Article : Google Scholar : PubMed/NCBI | |
|
Bossu P, Ciaramella A, Moro ML, Bellincampi L, Bernardini S, Federici G, Trequattrini A, Macciardi F, Spoletini I, Di Iulio F, et al: Interleukin 18 gene polymorphisms predict risk and outcome of Alzheimer's disease. J Neurol Neurosurg Psychiatry. 78:807–811. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Gasser T: Molecular pathogenesis of Parkinson disease: Insights from genetic studies. Expert Rev Mol Med. 11:e222009. View Article : Google Scholar : PubMed/NCBI | |
|
Braak H, Del Tredici K, Rub U, de Vos RA, Jansen EN and Braak E: Staging of brain pathology related to sporadic Parkinson's disease. Neurobiol Aging. 24:197–211. 2003. View Article : Google Scholar | |
|
Han C, Liu Y, Dai R, Ismail N, Su W and Li B: Ferroptosis and its potential role in human diseases. Front Pharmacol. 11:2392020. View Article : Google Scholar : PubMed/NCBI | |
|
Panicker N, Ge P, Dawson VL and Dawson TM: The cell biology of Parkinson's disease. J Cell Biol. 220:e2020120952021. View Article : Google Scholar : PubMed/NCBI | |
|
Liu G, Zhang C, Yin J, Li X, Cheng F, Li Y, Yang H, Uéda K, Chan P and Yu S: alpha-Synuclein is differentially expressed in mitochondria from different rat brain regions and dose-dependently down-regulates complex I activity. Neurosci Lett. 454:187–192. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Thomas RR, Keeney PM and Bennett JP: Impaired complex-I mitochondrial biogenesis in Parkinson disease frontal cortex. J Parkinsons Dis. 2:67–76. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Bender A, Krishnan KJ, Morris CM, Taylor GA, Reeve AK, Perry RH, Jaros E, Hersheson JS, Betts J, Klopstock T, et al: High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat Genet. 38:515–517. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Valente EM, Bentivoglio AR, Dixon PH, Ferraris A, Ialongo T, Frontali M, Albanese A and Wood NW: Localization of a novel locus for autosomal recessive early-onset parkinsonism, PARK6, on human chromosome 1p35-p36. Am J Hum Genet. 68:895–900. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Fan Z, Pan YT, Zhang ZY, Yang H, Yu SY, Zheng Y, Ma JH and Wang XM: Systemic activation of NLRP3 inflammasome and plasma α-synuclein levels are correlated with motor severity and progression in Parkinson's disease. J Neuroinflammation. 17:112020. View Article : Google Scholar | |
|
Sliter DA, Martinez J, Hao L, Chen X, Sun N, Fischer TD, Burman JL, Li Y, Zhang Z and Narendra DP: Parkin and PINK1 mitigate STING-induced inflammation. Nature. 561:258–262. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Hou X, Fiesel FC, Truban D, Casey MC, Lin WL, Soto AI, Tacik P, Rousseau LG, Diehl NN, Heckman MG, et al: Age- and disease-dependent increase of the mitophagy marker phospho-ubiquitin in normal aging and Lewy body disease. Autophagy. 14:1404–1418. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Di Maio R, Barrett PJ, Hoffman EK, Barrett CW, Zharikov A, Borah A, Hu X, McCoy J, Chu CT, Burton EA, et al: α-synuclein binds to TOM20 and inhibits mitochondrial protein import in Parkinson's disease. Sci Transl Med. 8:342ra3782016. View Article : Google Scholar | |
|
Vicario M, Cieri D, Vallese F, Catoni C, Barazzuol L, Berto P, Grinzato A, Barbieri L, Brini M and Calì T: A split-GFP tool reveals differences in the sub-mitochondrial distribution of wt and mutant alpha-synuclein. Cell Death Dis. 10:8572019. View Article : Google Scholar : PubMed/NCBI | |
|
Devi L, Raghavendran V, Prabhu BM, Avadhani NG and Anandatheerthavarada HK: Mitochondrial import and accumulation of alpha-synuclein impair complex I in human dopaminergic neuronal cultures and Parkinson disease brain. J Biol Chem. 283:9089–9100. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Mahul-Mellier AL, Burtscher J, Maharjan N, Weerens L, Croisier M, Kuttler F, Leleu M, Knott GW and Lashuel HA: The process of Lewy body formation, rather than simply alpha-synuclein fibrillization, is one of the major drivers of neurodegeneration. Proc Natl Acad Sci USA. 117:4971–4982. 2020. View Article : Google Scholar | |
|
Hsieh CH, Shaltouki A, Gonzalez AE, da Cruz AB, Burbulla LF, St Lawrence E, Schüle B, Krainc D, Palmer TD and Wang X: Functional impairment in miro degradation and mitophagy is a shared feature in familial and sporadic Parkinson's disease. Cell Stem Cell. 19:709–724. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Godena VK, Brookes-Hocking N, Moller A, Shaw G, Oswald M, Sancho RM, Miller CC, Whitworth AJ and De Vos KJ: Increasing microtubule acetylation rescues axonal transport and locomotor deficits caused by LRRK2 Roc-COR domain mutations. Nat Commun. 5:52452014. View Article : Google Scholar : PubMed/NCBI | |
|
Bonello F, Hassoun SM, Mouton-Liger F, Shin YS, Muscat A, Tesson C, Lesage S, Bear PM, Brice A, Krupp J, et al: LRRK2 impairs PINK1/Parkin-dependent mitophagy via its kinase activity: Pathologic insights into Parkinson's disease. Hum Mol Genet. 28:1645–1660. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Wang X, Chi J, Huang D, Ding L, Zhao X, Jiang L, Yu Y and Gao F: α-synuclein promotes progression of Parkinson's disease by upregulating autophagy signaling pathway to activate NLRP3 inflammasome. Exp Ther Med. 19:931–938. 2020.PubMed/NCBI | |
|
Sarkar S, Malovic E, Harishchandra DS, Ghaisas S, Panicker N, Charli A, Palanisamy BN, Rokad D, Jin H, Anantharam V, et al: Mitochondrial impairment in microglia amplifies NLRP3 inflammasome proinflammatory signaling in cell culture and animal models of Parkinson's disease. NPJ Parkinsons Dis. 3:302017. View Article : Google Scholar : PubMed/NCBI | |
|
Mouton-Liger F, Rosazza T, Sepulveda-Diaz J, Ieang A, Hassoun SM, Claire E, Mangone G, Brice A, Michel PP, Corvol JC and Corti O: Parkin deficiency modulates NLRP3 inflammasome activation by attenuating an A20-dependent negative feedback loop. Glia. 66:1736–1751. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Zhang C, Zhao M, Wang B, Su Z, Guo B, Qin L, Zhang W and Zheng R: The Nrf2-NLRP3-caspase-1 axis mediates the neuroprotective effects of celastrol in Parkinson's disease. Redox Biol. 47:1021342021. View Article : Google Scholar : PubMed/NCBI | |
|
Faissner S, Plemel JR, Gold R and Yong VW: Progressive multiple sclerosis: From pathophysiology to therapeutic strategies. Nat Rev Drug Discov. 18:905–922. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Barcelos IP, Troxell RM and Graves JS: Mitochondrial dysfunction and multiple sclerosis. Biology (Basel). 8:372019.PubMed/NCBI | |
|
Steinman L: Multiple sclerosis: A two-stage disease. Nat Immunol. 2:762–764. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Lassmann H, van Horssen J and Mahad D: Progressive multiple sclerosis: Pathology and pathogenesis. Nat Rev Neurol. 8:647–656. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Lopez-Domenech G and Kittler JT: Mitochondrial regulation of local supply of energy in neurons. Curr Opin Neurobiol. 81:1027472023. View Article : Google Scholar : PubMed/NCBI | |
|
Touil H, Li R, Zuroff L, Moore CS, Healy L, Cignarella F, Piccio L, Ludwin S, Prat A, Gommerman J, et al: Cross-talk between B cells, microglia and macrophages, and implications to central nervous system compartmentalized inflammation and progressive multiple sclerosis. EBioMedicine. 96:1047892023. View Article : Google Scholar : PubMed/NCBI | |
|
Campbell GR, Ziabreva I, Reeve AK, Krishnan KJ, Reynolds R, Howell O, Lassmann H, Turnbull DM and Mahad DJ: Mitochondrial DNA deletions and neurodegeneration in multiple sclerosis. Ann Neurol. 69:481–492. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Witte ME, Nijland PG, Drexhage JA, Gerritsen W, Geerts D, van Het Hof B, Reijerkerk A, de Vries HE, van der Valk P and van Horssen J: Reduced expression of PGC-1alpha partly underlies mitochondrial changes and correlates with neuronal loss in multiple sclerosis cortex. Acta Neuropathol. 125:231–243. 2013. View Article : Google Scholar | |
|
Haider L, Fischer MT, Frischer JM, Bauer J, Höftberger R, Botond G, Esterbauer H, Binder CJ, Witztum JL and Lassmann H: Oxidative damage in multiple sclerosis lesions. Brain. 134:1914–1924. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Dziedzic T, Metz I, Dallenga T, König FB, Müller S, Stadelmann C and Brück W: Wallerian degeneration: A major component of early axonal pathology in multiple sclerosis. Brain Pathol. 20:976–985. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Madsen PM, Pinto M, Patel S, McCarthy S, Gao H, Taherian M, Karmally S, Pereira CV, Dvoriantchikova G, Ivanov D, et al: Mitochondrial DNA double-strand breaks in oligodendrocytes cause demyelination, axonal injury, and CNS inflammation. J Neurosci. 37:10185–10199. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Singhal NK, Alkhayer K, Shelestak J, Clements R, Freeman E and McDonough J: Erythropoietin upregulates brain hemoglobin expression and supports neuronal mitochondrial activity. Mol Neurobiol. 55:8051–8058. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Aboul-Enein F, Krssak M, Hoftberger R, Prayer D and Kristoferitsch W: Reduced NAA-levels in the NAWM of patients with MS is a feature of progression. A study with quantitative magnetic resonance spectroscopy at 3 tesla. PLoS One. 5:e116252010. View Article : Google Scholar : PubMed/NCBI | |
|
Dominicis A, Del Giovane A, Torreggiani M, Recchia AD, Ciccarone F, Ciriolo MR and Ragnini-Wilson A: N-Acetylaspartate drives oligodendroglial differentiation via histone deacetylase activation. Cells. 12:18612023. View Article : Google Scholar : PubMed/NCBI | |
|
Kadowaki A and Quintana FJ: The NLRP3 inflammasome in progressive multiple sclerosis. Brain. 143:1286–1288. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Ferecsko AS, Smallwood MJ, Moore A, Liddle C, Newcombe J, Holley J, Whatmore J, Gutowski NJ and Eggleton P: STING-triggered CNS inflammation in human neurodegenerative diseases. Biomedicines. 11:13752023. View Article : Google Scholar : PubMed/NCBI | |
|
Thijs RD, Surges R, O'Brien TJ and Sander JW: Epilepsy in adults. Lancet. 393:689–701. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Fabisiak T and Patel M: Crosstalk between neuroinflammation and oxidative stress in epilepsy. Front Cell Dev Biol. 10:9769532022. View Article : Google Scholar : PubMed/NCBI | |
|
Loscher W and Klein P: The pharmacology and clinical efficacy of antiseizure medications: From bromide salts to cenobamate and beyond. CNS Drugs. 35:935–963. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Pearson-Smith JN and Patel M: Metabolic dysfunction and oxidative stress in epilepsy. Int J Mol Sci. 18:23652017. View Article : Google Scholar : PubMed/NCBI | |
|
Geronzi U, Lotti F and Grosso S: Oxidative stress in epilepsy. Expert Rev Neurother. 18:427–434. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Terrone G, Balosso S, Pauletti A, Ravizza T and Vezzani A: Inflammation and reactive oxygen species as disease modifiers in epilepsy. Neuropharmacology. 167:1077422020. View Article : Google Scholar | |
|
Ahras-Sifi N and Laraba-Djebari F: Immunomodulatory and protective effects of interleukin-4 on the neuropathological alterations induced by a potassium channel blocker. J Neuroimmunol. 355:5775492021. View Article : Google Scholar : PubMed/NCBI | |
|
Rahman S: Pathophysiology of mitochondrial disease causing epilepsy and status epilepticus. Epilepsy Behav. 49:71–75. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Liang LP, Waldbaum S, Rowley S, Huang TT, Day BJ and Patel M: Mitochondrial oxidative stress and epilepsy in SOD2 deficient mice: Attenuation by a lipophilic metalloporphyrin. Neurobiol Dis. 45:1068–1076. 2012. View Article : Google Scholar : | |
|
Fulton RE, Pearson-Smith JN, Huynh CQ, Fabisiak T, Liang LP, Aivazidis S, High BA, Buscaglia G, Corrigan T, Valdez R, et al: Neuron-specific mitochondrial oxidative stress results in epilepsy, glucose dysregulation and a striking astrocyte response. Neurobiol Dis. 158:1054702021. View Article : Google Scholar : PubMed/NCBI | |
|
Zhang S, Chen F, Zhai F and Liang S: Role of HMGB1/TLR4 and IL-1β/IL-1R1 signaling pathways in epilepsy. Front Neurol. 13:9042252022. View Article : Google Scholar | |
|
Kim JE and Kang TC: Differential roles of mitochondrial translocation of active caspase-3 and HMGB1 in neuronal death induced by status epilepticus. Front Cell Neurosci. 12:3012018. View Article : Google Scholar : PubMed/NCBI | |
|
Hyun HW, Ko AR and Kang TC: Mitochondrial translocation of high mobility group box 1 facilitates LIM kinase 2-mediated programmed necrotic neuronal death. Front Cell Neurosci. 10:992016. View Article : Google Scholar : PubMed/NCBI | |
|
Kim JE, Park H, Kim TH and Kang TC: LONP1 regulates mitochondrial accumulations of HMGB1 and Caspase-3 in CA1 and PV neurons following status epilepticus. Int J Mol Sci. 22:22752021. View Article : Google Scholar : PubMed/NCBI | |
|
Pauletti A, Terrone G, Shekh-Ahmad T, Salamone A, Ravizza T, Rizzi M, Pastore A, Pascente R, Liang LP, Villa BR, et al: Targeting oxidative stress improves disease outcomes in a rat model of acquired epilepsy. Brain. 142:e392019. View Article : Google Scholar : PubMed/NCBI | |
|
Kumar P, Osahon OW and Sekhar RV: GlyNAC (Glycine and N-Acetylcysteine) supplementation in mice increases length of life by correcting glutathione deficiency, oxidative stress, mitochondrial dysfunction, abnormalities in mitophagy and nutrient sensing, and genomic damage. Nutrients. 14:11142022. View Article : Google Scholar : PubMed/NCBI | |
|
Mohseni-Moghaddam P, Roghani M, Khaleghzadeh-Ahangar H, Sadr SS and Sala C: A literature overview on epilepsy and inflammasome activation. Brain Res Bull. 172:229–235. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Abais JM, Zhang C, Xia M, Liu Q, Gehr TW, Boini KM and Li PL: NADPH oxidase-mediated triggering of inflammasome activation in mouse podocytes and glomeruli during hyperhomocysteinemia. Antioxid Redox Signal. 18:1537–1548. 2013. View Article : Google Scholar : | |
|
Subramanian N, Natarajan K, Clatworthy MR, Wang Z and Germain RN: The adaptor MAVS promotes NLRP3 mitochondrial localization and inflammasome activation. Cell. 153:348–361. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Zhou R, Yazdi AS, Menu P and Tschopp J: A role for mitochondria in NLRP3 inflammasome activation. Nature. 469:221–225. 2011. View Article : Google Scholar | |
|
Rong S, Wan D, Fan Y, Liu S, Sun K, Huo J, Zhang P, Li X, Xie X, Wang F and Sun T: Amentoflavone affects epileptogenesis and exerts neuroprotective effects by inhibiting NLRP3 inflammasome. Front Pharmacol. 10:8562019. View Article : Google Scholar : PubMed/NCBI | |
|
Liu Z, Xian H, Ye X, Chen J, Ma Y and Huang W: Increased levels of NLRP3 in children with febrile seizures. Brain Dev. 42:336–341. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
de Brito Toscano EC, Vieira EL, Dias BB, Caliari MV, Gonçalves AP, Giannetti AV, Siqueira JM, Suemoto CK, Leite RE, Nitrini R, et al: NLRP3 and NLRP1 inflammasomes are up-regulated in patients with mesial temporal lobe epilepsy and may contribute to overexpression of caspase-1 and IL-β in sclerotic hippocampi. Brain Res. 1752:1472302021. View Article : Google Scholar | |
|
Tan CC, Zhang JG, Tan MS, Chen H, Meng DW, Jiang T, Meng XF, Li Y, Sun Z, Li MM, et al: NLRP1 inflammasome is activated in patients with medial temporal lobe epilepsy and contributes to neuronal pyroptosis in amygdala kindling-induced rat model. J Neuroinflammation. 12:182015. View Article : Google Scholar : PubMed/NCBI | |
|
Samidurai M, Tarale P, Janarthanam C, Estrada CG, Gordon R, Zenitsky G, Jin H, Anantharam V, Kanthasamy AG and Kanthasamy A: tumor necrosis factor-like weak inducer of apoptosis (TWEAK) enhances activation of STAT3/NLRC4 inflammasome signaling axis through PKCdelta in AStrocytes: Implications for Parkinson's disease. Cells. 9:18312020. View Article : Google Scholar | |
|
Zadori D, Klivenyi P, Szalardy L, Fulop F, Toldi J and Vecsei L: Mitochondrial disturbances, excitotoxicity, neuroinflammation and kynurenines: Novel therapeutic strategies for neurodegenerative disorders. J Neurol Sci. 322:187–191. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Prasuhn J, Davis RL and Kumar KR: Targeting mitochondrial impairment in Parkinson's disease: Challenges and opportunities. Front Cell Dev Biol. 8:6154612020. View Article : Google Scholar | |
|
Vos M, Esposito G, Edirisinghe JN, Vilain S, Haddad DM, Slabbaert JR, Van Meensel S, Schaap O, De Strooper B, Meganathan R, et al: Vitamin K2 is a mitochondrial electron carrier that rescues pink1 deficiency. Science. 336:1306–1310. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Bronzuoli MR, Iacomino A, Steardo L and Scuderi C: Targeting neuroinflammation in Alzheimer's disease. J Inflamm Res. 9:199–208. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Fu AK, Hung KW, Yuen MY, Zhou X, Mak DS, Chan IC, Cheung TH, Zhang B, Fu WY, Liew FY and Ip NY: IL-33 ameliorates Alzheimer's disease-like pathology and cognitive decline. Proc Natl Acad Sci USA. 113:E2705–E2713. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Wang X, Sun G, Feng T, Zhang J, Huang X, Wang T, Xie Z, Chu X, Yang J, Wang H, et al: Sodium oligomannate therapeutically remodels gut microbiota and suppresses gut bacterial amino acids-shaped neuroinflammation to inhibit Alzheimer's disease progression. Cell Res. 29:787–803. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Regen F, Hellmann-Regen J, Costantini E and Reale M: Neuroinflammation and Alzheimer's disease: Implications for microglial activation. Curr Alzheimer Res. 14:1140–1148. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
La Torre ME, Cianciulli A, Monda V, Monda M, Filannino FM, Antonucci L, Valenzano A, Cibelli G, Porro C, Messina G, et al: α-Tocopherol protects lipopolysaccharide-activated BV2 microglia. Molecules. 28:33402023. View Article : Google Scholar | |
|
Shabab T, Khanabdali R, Moghadamtousi SZ, Kadir HA and Mohan G: Neuroinflammation pathways: A general review. Int J Neurosci. 127:624–633. 2017. View Article : Google Scholar | |
|
Heger LM, Wise RM, Hees JT, Harbauer AB and Burbulla LF: Mitochondrial phenotypes in Parkinson's diseases-a focus on human iPSC-derived dopaminergic neurons. Cells. 10:34362021. View Article : Google Scholar : PubMed/NCBI | |
|
Yu G, Wang Y and Zhao J: Inhibitory effect of mitoquinone against the α-synuclein fibrillation and relevant neurotoxicity: Possible role in inhibition of Parkinson's disease. Biol Chem. 403:253–263. 2022. View Article : Google Scholar | |
|
Snow BJ, Rolfe FL, Lockhart MM, Frampton CM, O'Sullivan JD, Fung V, Smith RA, Murphy MP and Taylor KM; Protect Study Group: A double-blind, placebo-controlled study to assess the mitochondria-targeted antioxidant MitoQ as a disease-modifying therapy in Parkinson's disease. Mov Disord. 25:1670–1674. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Lehmann S, Loh SH and Martins LM: Enhancing NAD(+) salvage metabolism is neuroprotective in a PINK1 model of Parkinson's disease. Biol Open. 6:141–147. 2017. | |
|
Monti DA, Zabrecky G, Kremens D, Liang TW, Wintering NA, Bazzan AJ, Zhong L, Bowens BK, Chervoneva I, Intenzo C and Newberg AB: N-Acetyl cysteine is associated with dopaminergic improvement in Parkinson's disease. Clin Pharmacol Ther. 106:884–890. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Sathe AG, Tuite P, Chen C, Ma Y, Chen W, Cloyd J, Low WC, Steer CJ, Lee BY, Zhu XH and Coles LD: Pharmacokinetics, safety, and tolerability of orally administered ursodeoxycholic acid in patients with Parkinson's disease-a pilot study. J Clin Pharmacol. 60:744–750. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Bell SM, Barnes K, Clemmens H, Al-Rafiah AR, Al-Ofi EA, Leech V, Bandmann O, Shaw PJ, Blackburn DJ, Ferraiuolo L and Mortiboys H: Ursodeoxycholic acid improves mitochondrial function and redistributes Drp1 in fibroblasts from patients with either sporadic or familial Alzheimer's Disease. J Mol Biol. 430:3942–3953. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Lin MW, Lin CC, Chen YH, Yang HB and Hung SY: Celastrol inhibits dopaminergic neuronal death of Parkinson's disease through activating mitophagy. Antioxidants (Basel). 9:372019. View Article : Google Scholar | |
|
Bido S, Soria FN, Fan RZ, Bezard E and Tieu K: Mitochondrial division inhibitor-1 is neuroprotective in the A53T-α-synuclein rat model of Parkinson's disease. Sci Rep. 7:74952017. View Article : Google Scholar | |
|
Curtis WM, Seeds WA, Mattson MP and Bradshaw PC: NADPH and mitochondrial quality control as targets for a circadian-based fasting and exercise therapy for the treatment of Parkinson's disease. Cells. 11:24162022. View Article : Google Scholar : PubMed/NCBI | |
|
Ferreira AFF, Binda KH, Singulani MP, Pereira CPM, Ferrari GD, Alberici LC, Real CC and Britto LR: Physical exercise protects against mitochondria alterations in the 6-hidroxydopamine rat model of Parkinson's disease. Behav Brain Res. 387:1126072020. View Article : Google Scholar : PubMed/NCBI | |
|
Kappos L, Bar-Or A, Cree BAC, Fox RJ, Giovannoni G, Gold R, Vermersch P, Arnold DL, Arnould S, Scherz T, et al: Siponimod versus placebo in secondary progressive multiple sclerosis (EXPAND): A double-blind, randomised, phase 3 study. Lancet. 391:1263–1273. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Montalban X, Hauser SL, Kappos L, Arnold DL, Bar-Or A, Comi G, de Seze J, Giovannoni G, Hartung HP, Hemmer B, et al: Ocrelizumab versus placebo in primary progressive multiple sclerosis. N Engl J Med. 376:209–220. 2017. View Article : Google Scholar | |
|
Lublin F, Miller DH, Freedman MS, Cree BAC, Wolinsky JS, Weiner H, Lubetzki C, Hartung HP, Montalban X, Uitdehaag BMJ, et al: Oral fingolimod in primary progressive multiple sclerosis (INFORMS): A phase 3, randomised, double-blind, placebo-controlled trial. Lancet. 387:1075–1084. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Waxman SG: Axonal conduction and injury in multiple sclerosis: The role of sodium channels. Nat Rev Neurosci. 7:932–941. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Picone P and Nuzzo D: Promising treatment for multiple sclerosis: Mitochondrial transplantation. Int J Mol Sci. 23:22452022. View Article : Google Scholar : PubMed/NCBI | |
|
Moos WH, Faller DV, Glavas IP, Kanara I, Kodukula K, Pernokas J, Pernokas M, Pinkert CA, Powers WR, Sampani K, et al: Epilepsy: Mitochondrial connections to the 'Sacred' disease. Mitochondrion. 72:84–101. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Silvestro S, Mammana S, Cavalli E, Bramanti P and Mazzon E: Use of cannabidiol in the treatment of epilepsy: Efficacy and security in clinical trials. Molecules. 24:14592019. View Article : Google Scholar : PubMed/NCBI | |
|
Ramirez A, Old W, Selwood DL and Liu X: Cannabidiol activates PINK1-Parkin-dependent mitophagy and mitochondrial-derived vesicles. Eur J Cell Biol. 101:1511852022. View Article : Google Scholar : | |
|
Bhunia S, Kolishetti N, Arias AY, Vashist A and Nair M: Cannabidiol for neurodegenerative disorders: A comprehensive review. Front Pharmacol. 13:9897172022. View Article : Google Scholar : PubMed/NCBI | |
|
Britch SC, Babalonis S and Walsh SL: Cannabidiol: Pharmacology and therapeutic targets. Psychopharmacology (Berl). 238:9–28. 2021. View Article : Google Scholar | |
|
Aledo-Serrano A, Hariramani R, Gonzalez-Martinez A, Álvarez-Troncoso J, Toledano R, Bayat A, Garcia-Morales I, Becerra JL, Villegas-Martínez I, Beltran-Corbellini A and Gil-Nagel A: Anakinra and tocilizumab in the chronic phase of febrile infection-related epilepsy syndrome (FIRES): Effectiveness and safety from a case-series. Seizure. 100:51–55. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Stienen MN, Haghikia A, Dambach H, Thöne J, Wiemann M, Gold R, Chan A, Dermietzel R, Faustmann PM, Hinkerohe D and Prochnow N: Anti-inflammatory effects of the anticonvulsant drug levetiracetam on electrophysiological properties of astroglia are mediated via TGFβ1 regulation. Br J Pharmacol. 162:491–507. 2011. View Article : Google Scholar : | |
|
Stockburger C, Miano D, Baeumlisberger M, Pallas T, Arrey TN, Karas M, Friedland K and Müller WE: A mitochondrial role of sv2a protein in aging and Alzheimer's disease: Studies with levetiracetam. J Alzheimers Dis. 50:201–215. 2016. View Article : Google Scholar | |
|
Yang N, Guan QW, Chen FH, Xia QX, Yin XX, Zhou HH and Mao XY: Antioxidants targeting mitochondrial oxidative stress: Promising neuroprotectants for epilepsy. Oxid Med Cell Longev. 2020:66871852020. View Article : Google Scholar : PubMed/NCBI | |
|
Fields M, Marcuzzi A, Gonelli A, Celeghini C, Maximova N and Rimondi E: Mitochondria-Targeted antioxidants, an innovative class of antioxidant compounds for neurodegenerative diseases: Perspectives and limitations. Int J Mol Sci. 24:37392023. View Article : Google Scholar : PubMed/NCBI | |
|
Huenchuguala S and Segura-Aguilar J: Single-neuron neurodegeneration as a degenerative model for Parkinson's disease. Neural Regen Res. 19:529–535. 2024. View Article : Google Scholar | |
|
Leitao-Rocha A, Guedes-Dias P, Pinho BR and Oliveira JM: Trends in mitochondrial therapeutics for neurological disease. Curr Med Chem. 22:2458–2467. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Yoshinaga N and Numata K: Rational designs at the forefront of mitochondria-targeted gene delivery: Recent progress and future perspectives. ACS Biomater Sci Eng. 8:348–359. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Silva-Pinheiro P and Minczuk M: The potential of mitochondrial genome engineering. Nat Rev Genet. 23:199–214. 2022. View Article : Google Scholar |









