



mRNA vaccine development and applications: A special focus on tumors (Review)
- Authors:
- Published online on: July 9, 2024 https://doi.org/10.3892/ijo.2024.5669
- Article Number: 81
-
Copyright: © Chen et al. This is an open access article distributed under the terms of Creative Commons Attribution License.
Abstract
1. Introduction
Cancer remains a serious threat to human health and the incidence rates continue to rise. In recent years, the number of new cases and cancer-associated deaths has shown a significant upward trend (1-3). The occurrence of cancer is a multi-factor, multi-step and complex process, and its etiology has not been fully clarified; however, it is well acknowledged that it is closely related to various factors, including smoking, infection, occupational exposures, environmental pollution, an unhealthy diet and genetic factors (4-8). With the rapid advances in life sciences and technology, a more in-depth understanding of tumors has been gained. The search for more effective treatments to improve the survival rates of patients with cancer remains a central topic of research in the medical industry.
mRNA vaccines are a type of nucleic acid preparation that can transduce gene sequences of exogenous target antigens into cells via a specific delivery system by means of transcription and translation, and the transcribed proteins stimulate the body to produce specific immunological responses, thus enabling the body to obtain immune protection (9). mRNA vaccines are the new generation of vaccines after inactivated vaccines, live attenuated vaccines, subunit vaccines and viral vector vaccines. With continuous improvements in RNA-based technologies, this innovative treatment model is predicted to supplant traditional vaccines and may have the ability to address the issues associated with small molecules and antibody therapy. In addition to being used in infectious diseases, mRNA vaccines also provide more effective and longer-lasting treatment opportunities for diseases including cancer, rare diseases and nervous system diseases (10). mRNA vaccines not only cover a variety of tumor antigens at one time but also activate a wider range of T-cell responses through the simultaneous delivery of human leukocyte antigen (HLA)-I and HLA-II molecules, resulting in a more comprehensive multi-angle attack. Thus, mRNA vaccines have significant anti-cancer potential. Currently, there are dozens of clinical trials related to mRNA tumor vaccines around the world, which include (but are not limited to) melanoma, pancreatic cancer and colorectal cancer, and in certain trials, mRNA vaccines are combined with other tumor-immunity drugs (11-13). The recently developed mRNA-4157 vaccine, which has been declared a 'Breakthrough Therapy Designation' by the US Food and Drug Administration (FDA) and a 'Priority Medicines' Programme by the European Medicines Agency, is expected to be the first mRNA tumor vaccine on the market (14).
The present article reviews the current treatment methods for cancer as well as the development and current status of mRNA vaccines. On this basis, the value of mRNA vaccines in tumor treatment is also discussed.
2. Treatment of cancer
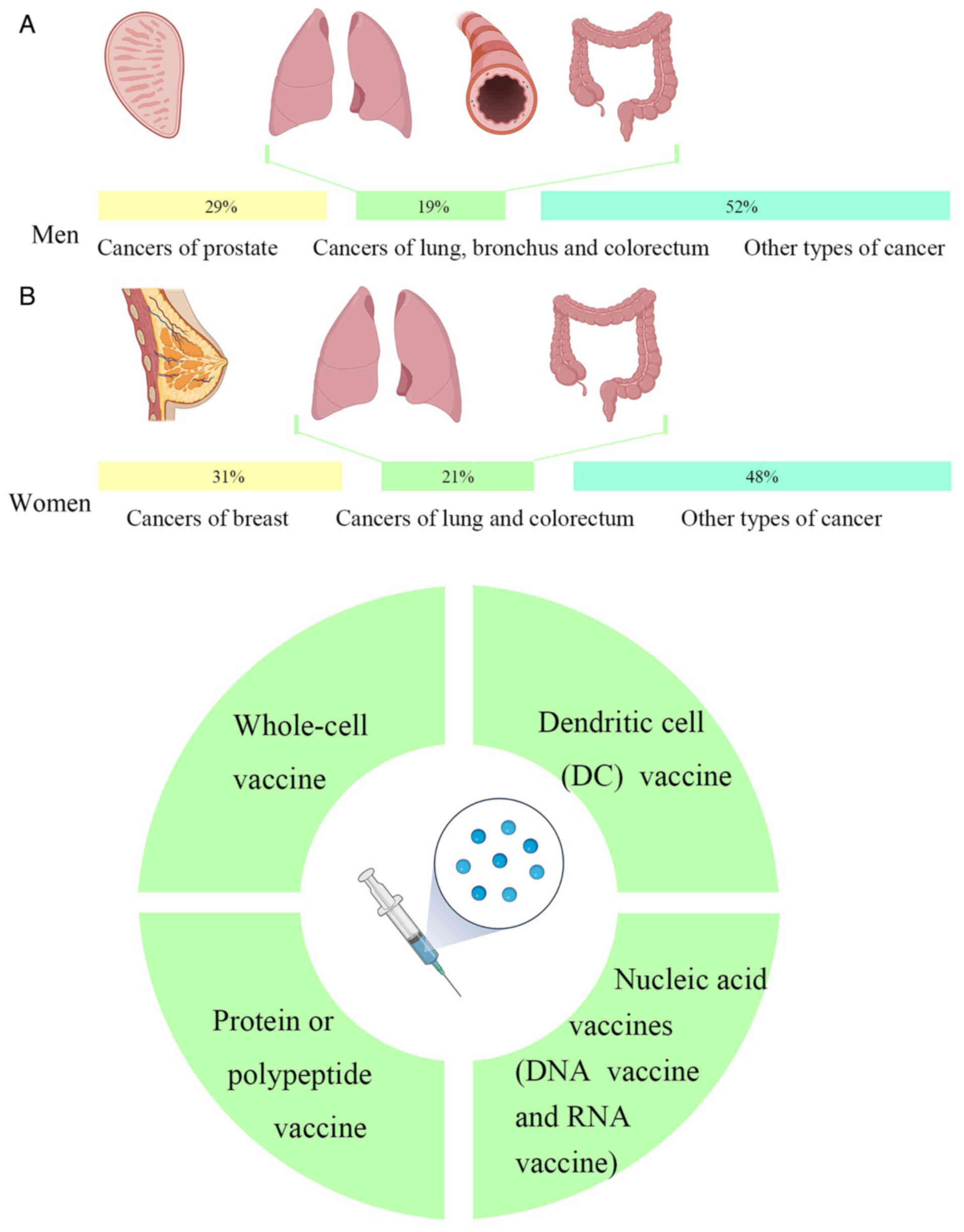
Cancer is a major global public health problem. According to the latest global cancer data released by the American Cancer Society, it is estimated that there were 1,958,310 new cancer cases and 609,820 deaths in the US in 2023. Amongst the different types of cancer, cancers of the prostate, lung, bronchus and colorectum account for almost half (48%) of all cases in men, with prostate cancer alone accounting for 29% of all diagnoses. For women, breast, lung and colorectal cancers account for 52% of all newly diagnosed cancers, with breast cancer (BCa) alone accounting for 31% of cancers in women. Of the projected deaths from cancer, the highest number of deaths were from lung, prostate and colorectal cancers in men and from lung, breast and colorectal cancers in women (Fig. 1A) (15).
Due to the continuous progress of medical approaches, there are an ever-increasing number of methods for treating cancer. According to the type and stage of cancer and the specific conditions of a patient, choosing the appropriate treatment can prolong the survival time and improve the quality of life to a significant extent. The methods of cancer treatment primarily include surgery, chemotherapy, radiotherapy and immunotherapy. However, a single treatment is insufficient for the treatment of cancer and the simple addition of a therapeutic regimen cannot achieve the goal. Thus, a reasonable and comprehensive use of a variety of methods is required to better control and eliminate the tumor; that is, a current comprehensive multidisciplinary approach should be adopted where possible (16).
With the development of vaccinations being promoted by emerging innovations of the digital age, vaccinating a patient with individual tumor mutations may become the first truly personalized treatment for cancer (17). All types of vaccines that can stimulate the body to produce agents that counteract pathogenic factors for tumorigenesis or activate specific immunity in order to counteract the immune escape effect of cancers and treat tumors can be called tumor vaccines (18). Prophylactic vaccines, which are similar to traditional vaccines, primarily target pathogenic microorganisms directly related to cancer, such as Human papillomavirus (HPV) and hepatitis B virus (HBV). In addition, there are therapeutic vaccines for patients who already have cancer and these promote the establishment of a lasting immune memory and a long-term anti-tumor response by inducing immune responses primarily against 'autoantigens' present in tumors (19).
At present, the primary types of tumor vaccines are as follows: i) Whole-cell vaccines; ii) dendritic cell (DC) vaccines; iii) protein or polypeptide vaccines; and iv) nucleic acid vaccines, including DNA vaccines and RNA vaccines (Fig. 1B).
3. Overview of mRNA vaccines
History of mRNA vaccines
During the Coronavirus (CoV) disease 2019 (COVID-19) pandemic, mRNA vaccines gained attention as a cutting-edge technology used by several pharmaceutical companies to create vaccinations (20,21). Of note, the first potential vaccine to enter phase I clinical trials was an mRNA vaccine. Brenner et al (22) first identified mRNAs as an intermediary genetic element in the central nervous system in 1961. Malone et al (23) in 1989 discovered that mRNAs could be successfully transfected and expressed in a variety of eukaryotic cells when encased in a cationic lipid {N-[1-(2,3-dioleyloxy) propyl]-N,N,N-trimethylammonium chloride}, and this was when the notion of mRNA-based medications was first conceived. The 1990s saw a boom in mRNA vaccine research, with preclinical trials being applied to a range of illnesses, including infectious diseases and cancer. The first successful effort at mRNA in vivo expression was performed in 1990 when direct injection of in vitro-transcribed mRNA was sufficient to introduce the mRNA into mouse skeletal muscle cells (24). This demonstrated the viability of mRNA vaccines. In 1992, Jirikowski et al (25) discovered that injecting mRNAs encoding oxytocin and vasopressin into genetically mutated diabetic insipidus mice cured the condition within a few hours of injection for a short period of time. An in vitro-generated mRNA vaccine encoding the influenza virus's nucleoprotein to activate cytotoxic T-cells of mice was reported by Martinon et al (26) in 1993. The first researchers to find that DCs pulsed with mRNA was a viable approach to trigger T-cell responses were Boczkowski et al (27) in 1996. Zhou et al (28), in 1999, showed the potential of mRNA vaccines against cancers by directly injecting glycoprotein 100 mRNA encapsulated in hemagglutinin virus of Japan-liposomes into the spleen, which resulted in tumor growth restriction and a longer survival time in a mouse model of melanoma. The safety, effectiveness and industrial manufacturing ability of mRNA vaccines have improved over the last several decades as research has advanced and experimental procedures have improved (29). Currently, several mRNA vaccines are being tested in clinical settings or are readily accessible to combat cancer and several infectious diseases, including the Zika virus, cytomegalovirus, influenza virus, metapneumovirus and parainfluenza virus (30). In response to the need for a timely and efficient vaccination, there has been a surge in the research and development of nucleic acid-based vaccines since the worldwide severe acute respiratory syndrome (SARS) CoV-2 pandemic (21,31). Moreover, the market value of mRNA vaccines has grown to tens of billions of dollars, indicating a promising future for the development of mRNA-based medications, particularly mRNA vaccines (32) (Fig. 2).

Mechanism of mRNA vaccines
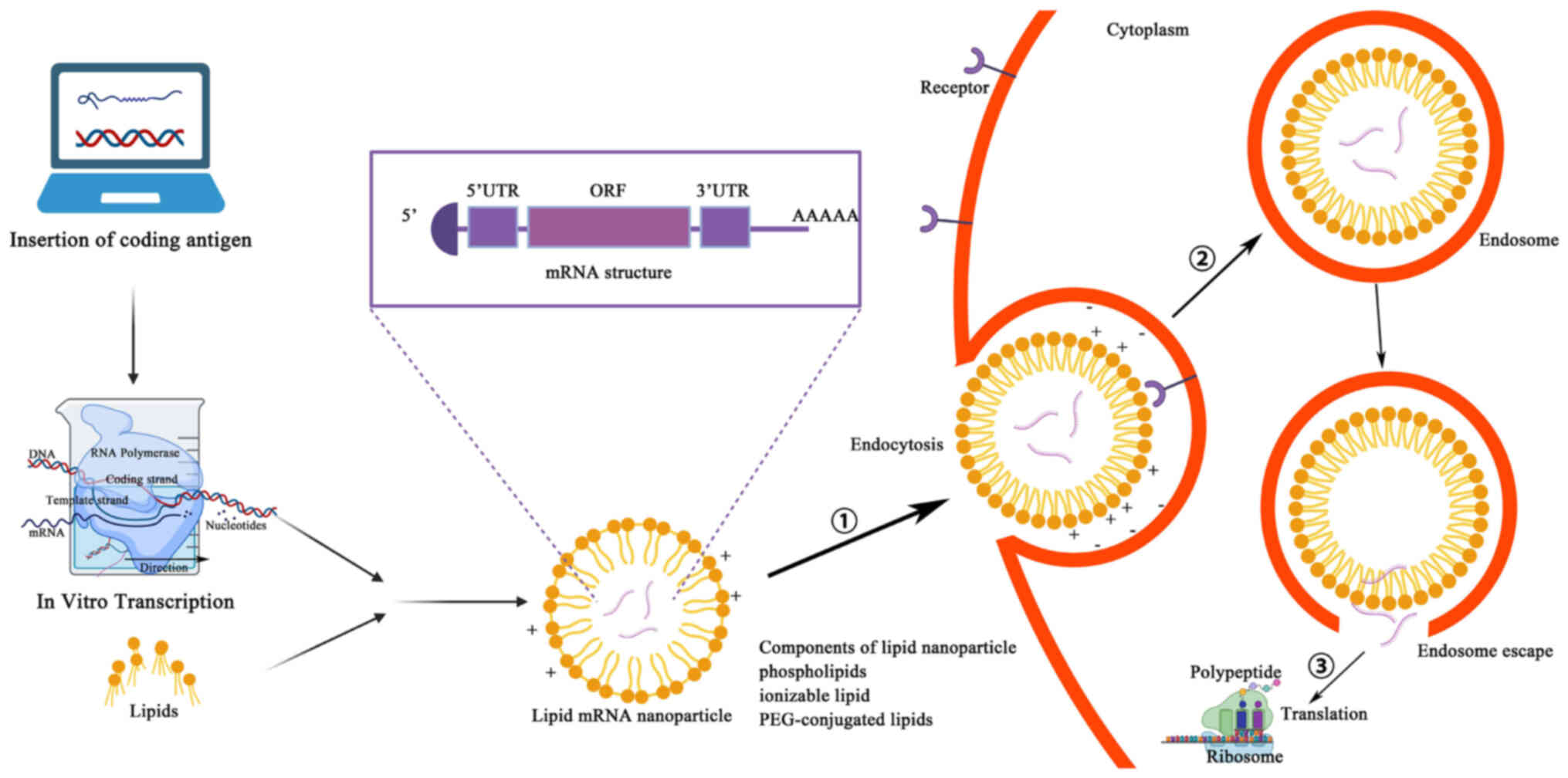
Manufacturing an mRNA vaccine involves several relatively complex steps. First, the desired mRNA sequence needs to be designed and synthesized; this sequence usually includes the nucleotide sequence encoding the target antigen or another therapeutic protein, and should also contain the appropriate 5' end cap and 3' end tail sequences to improve mRNA stability and translation efficiency (33). Through in vitro transcription, the corresponding mRNA is synthesized, with the in vitro transcription system typically consisting of RNA polymerase, buffer solution, nucleoside triphosphates, zinc ions and cofactors to facilitate transcription. Mixing the DNA template with the other components of the in vitro transcription system constitutes a reaction mixture containing all the necessary components for synthesizing mRNA. This reaction mixture is then placed into appropriate reaction tubes and subjected to an in vitro transcription reaction under the appropriate conditions. During this reaction, the RNA polymerase identifies the DNA template and uses it to synthesize mRNA (34). Synthetic mRNAs need to be purified and modified to remove impurities and improve stability and certain mRNA vaccines may need to be encapsulated in liposomes to improve their stability and promote cellular uptake in vivo. Liposomes are usually composed of constituents such as phospholipids and cholesterol, which can form tiny lipid vesicles in the aqueous phase with a lipid bilayer structure. Surfactants (such as polyethylene glycol) can be added to improve the stability and cellular uptake efficiency of liposomes. The synthesized mRNA is mixed with liposomes to form liposome-mRNA complexes. These complexes can be transfected into target cells by an endocytotic mechanism, whereby the cell membrane forms invaginations to carry the liposome-mRNA complexes inside the cell. The endocytosed vesicles encapsulate the liposome-mRNA complexes in endosomes, which in turn fuse with other organelles in the cytoplasm and release their contents into the cytoplasm. After the contents of the liposome-mRNA complex are released into the cytoplasm, the mRNA molecules are recognized and translated by intracellular ribosomes (35,36) (Fig. 3). A fraction of this foreign mRNA eludes destruction by ubiquitous RNase and is internalized by natural processes unique to individual cells (such as macropinocytosis in developing DCs) into the endosomal pathway, where it is released into the cytoplasm by the endosomes (37). Antigenic proteins are translated from the mRNA by the host cell's translation machinery. Signal peptides dictate the encoded protein's final location once it has been produced. These may be recombinantly created to target the protein to the appropriate cellular compartment inside the host cell, or they can be inherent to the original protein's sequence (37). As an alternative, this encoded protein may affect the function of nearby cells via its secretion, and it can also affect distant organs if it is released into the blood. Proteases in the cytoplasm break down antigenic proteins that are important to cells, creating antigenic peptide epitopes. The endoplasmic reticulum is where antigenic peptide epitopes are transferred, loaded onto class I molecules of the major histocompatibility complex (MHC I) and presented to CD8+ cytotoxic T-cells. The protein product has to be directed to MHC class II loading compartments in antigen-presenting cells (APCs) in order to enlist the assistance of cognate T-cells to induce a more powerful and longer-lasting immune response (10,37). This may be achieved by adding routing signal-encoding sequences to the mRNA. After the produced antigen peptide epitope is loaded onto the MHC II molecule, the antigen-specific CD4+ T-cell response is induced and the loaded MHC II peptide epitope complex emerges on the cell surface (33) (Fig. 4). Furthermore, via a process called cross-priming, foreign antigens that are taken up by DCs may also be processed and loaded onto MHC class I molecules (38). This allows for the design, in vitro production and delivery of any desired sequence to any kind of cell (39).
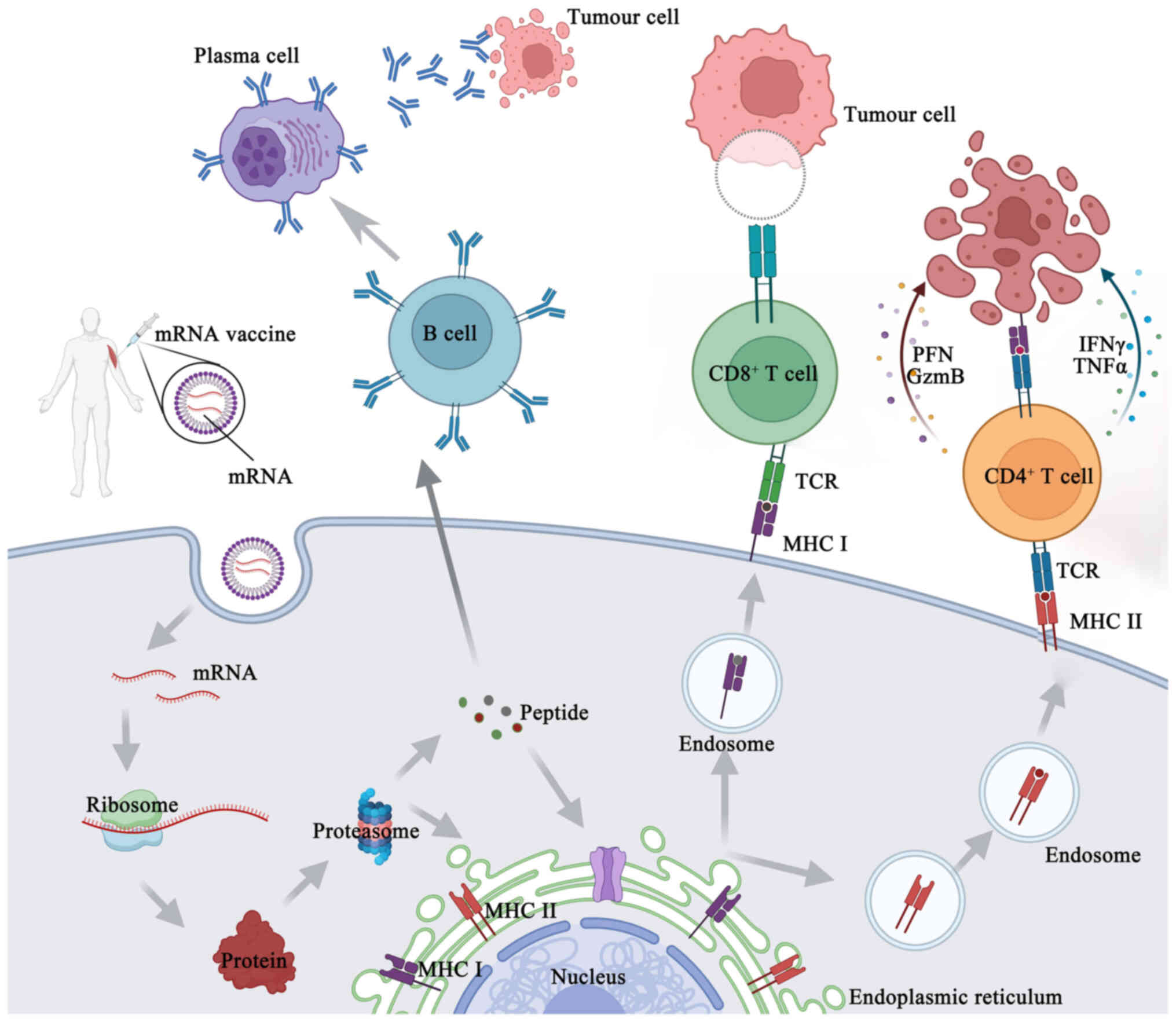
When mRNA functions as the vector of foreign genes, it may display certain characteristics similar to those of the mRNA virus due to its self-adjuvant impact. APCs can identify mRNAs prior to translation and this identification triggers the activation of pattern recognition receptors, including Toll-like receptor (TLR)3, TLR7 and TLR8 (40). Single-stranded and double-stranded (ds) RNA chains can be detected by TLR3, TLR7 and TLR8, while short and long filaments of dsRNA can be detected in the cytosol by retinoic acid-inducible gene-I (RIG-I) and melanoma differentiation-associated protein 5 (41,42). As a result, the type I interferon (IFN-I) pathway may be activated, chemokines and proinflammatory cytokines may be produced and APCs may be activated, all of which may trigger a robust adaptive response (43).
Advantages of mRNA vaccines
The mRNA tumor vaccine platform offers unique advantages over other cancer therapies. Different from traditional radiotherapy and chemotherapy, which are susceptible to drug resistance, mRNA vaccines allow for the simultaneous coverage of multiple antigens, thus overcoming vaccine resistance in cancer treatment. The combination of mRNA vaccines and immune checkpoint inhibitors (ICIs) can play a role in reversing the pathways that allow for resistance. Therefore, mRNA vaccines are expected to be a useful adjunct to ICIs and the combination of the two may allow for the implementation of precise and individualized treatment for patients (44). In addition, conventional cancer treatments can cause significant damage to normal human cells, whereas mRNA vaccines can target tumor cells more precisely. Furthermore, the transient nature of mRNA expression ensures that antigenic stimulation is temporary, which effectively reduces the possibility of chronic inflammation or autoimmune reactions (45). Compared with other tumor vaccine platforms, mRNA vaccines are highly immunogenic, stimulating a broader immune response in patients.
While mRNA tumor vaccines show clinical potential, their economic value is also noteworthy. As a novel therapy, mRNA vaccines have advantages over other cell and gene therapy products due to their convenience and low cost of production. Other gene/cell therapies already on the market, such as chimeric antibody receptor T-cell immunotherapy (CAR-T), are currently priced at millions of dollars, which poses a significant financial burden to patients and society (46,47). This is due to the fact that CAR-T requires a series of processes such as cell extraction, modification, in vitro culture and re-infusion for patients, and it is difficult to scale up the production, thus making it difficult to reduce the cost of a single product. By contrast, mRNA vaccines are directly transcribed in vitro. The synthesis and production steps for mRNA vaccines are standardized, allowing for the rapid and large-scale production of the product, which would result in a significant cost advantage of the product. Therefore, mRNA vaccines are expected to provide a boon to less economically privileged cancer patients and thus have significant economic potential in the current medical environment.
Given the several benefits, mRNA technology is a more appealing option than DNA or even conventional vaccinations. In vitro transcription (IVT) synthesis eliminates the need for cells and the regulatory barriers that go along with them, making mRNA production unquestionably easier, faster and cleaner than large-scale protein production and purification (48,49), in contrast to conventional vaccination approaches. The accuracy of antigen design allows for a quicker response to new risks of pandemics and epidemics in a timely and efficient manner (50). mRNA is precise because it only produces certain antigens and triggers a tailored immune response, in contrast to attenuated or inactivated vaccines. mRNA may trigger and/or promote an adaptive immune response by binding to pattern recognition receptors without the need for adjuvants, while subunit vaccines need adjuvants to elicit an immunological response. Nucleic acid vaccines are less constrained by HLA types and are more likely to elicit a wider T-cell response than peptide vaccines, as they can encode full-length tumor antigens, which allows APCs to cross-present or present multiple epitopes with both class I and II patient-specific HLA at the same time (49,51).
The following describes the ways in which mRNA vaccines are superior to DNA vaccines. The nuclear envelope barrier that prevents DNA vaccines from working is removed, as only mRNA has to be transcribed in the cytoplasm, while DNA must be translated in the nucleus (52). Consequently, both mitotic and non-mitotic cells may be successfully transfected with mRNA vaccines (53). Furthermore, mRNA is rarely integrated into genomic DNA and does not undergo insertion mutation. Third, since mRNA is rapidly broken down by biological processes and leaves no trace after 2-3 days, the production of the coded antigens is only temporary. The mRNA vaccine platform's flexibility is also useful for manufacturing, since it allows for standardization of production, as modifications to the encoded antigen do not alter the physical/chemical properties of the mRNA backbone. Fourth, there are fewer safety concerns regarding the presence of viral contaminants and cell-derived impurities than with other platforms, since production is based on an in vitro cell-free transcription process (39). Furthermore, RNA vaccinations often induce stronger immune responses than DNA vaccines. The reasons for this are not entirely clear, but some theories include low expression of DNA-sensing machinery, different expression of nucleic acid sensing pattern recognition receptors, inefficient delivery of DNA into human cells and the requirement that DNA crosses both cell and nuclear membranes and is transcribed in the nucleus in order to successfully transfect a cell (34,54).
Potential limitations of mRNA vaccines
Although the current success of the SARS-CoV-2 mRNA vaccine has fueled research into mRNA vaccines for cancer, there are still limitations to the application of mRNA vaccines in cancer, which are discussed in the following chapters.
Harsh preservation conditions
mRNA is an unstable molecule that can be easily cleaved by biological nucleases in the environment due to the presence of hydroxyl groups on the ribose. Stable activity can be maintained when it is cryopreserved, but the cost of storage is too high for economically underdeveloped regions (55). Carrier-based protectants such as lipid nanoparticles (LNPs) can only preserve mRNA vaccines for a maximum of 3 months (56). Cichlidin also protects mRNA from enzymatic degradation but may make it more difficult to translate, affecting the efficacy of the mRNA vaccine (57,58).
Relatively low anti-tumor effect when used alone
It is difficult to qualitatively improve the anti-tumor effect of mRNA vaccines alone due to their poor stability, the relatively short production cycle of the proteins, the heterogeneity of tumor cells and the fact that the immune microenvironment of advanced tumors is often highly immunosuppressive. As a result, clinical studies on cancer mRNA vaccines have shown limited progress (13).
Lack of long-term experimental findings
Clinical trials for cancer mRNA vaccines are currently active and generally in the early stages of development. Clinical trials with reported results, particularly for personalized cancer mRNA vaccines, are still limited and the therapeutic efficacy varies widely among patients. There is still a lack of results from long-term trials of mRNA vaccines in cancer, so the roadmap to their use as a routine treatment for cancer is long. It has been reported that mRNA vaccines may be associated with certain rare and fatal thrombotic events, so the safety of mRNA vaccine administration is also worth observing long-term (55). In summary, it is essential to monitor the long-term efficacy and side effects of mRNA vaccines in cancer therapy.
Regulatory challenges and ethical considerations
The development and production of mRNA vaccines for personalized neoantigens is directed at the genomic data of individuals, and as a result, the vaccines can pose regulatory and approval challenges.
Factors influencing mRNA vaccines
Despite all of the benefits of mRNA vaccines, their fundamental qualities nevertheless place certain restrictions on their use in the treatment of cancer. For example, the negative charge of both mRNA and the cell membrane makes mRNA distribution more challenging (34,37,59,60). Skin and blood contain extracellular ribonucleases, which readily break down mRNA (61). Given its inherent immunogenicity, mRNA may trigger innate immunity by activating a downstream interferon-related pathway. Paradoxically, this inherent immunogenicity promotes mRNA degradation, which lowers antigen expression even though it may be used as an adjuvant-like effect to increase the immune response (49). dsRNA contaminants are often found in mRNA in vitro transcription products. Type I IFN production may be enhanced by dsRNA, a simulant of RNA virus genome replication intermediates. This further restricts the translation of mRNA and strengthens the activation of innate immunity (62). Several techniques have been developed to advance the science of mRNA vaccines in response to these issues.
Delivery systems for mRNA vaccines
It is well established that mRNA has a half-life of ~7 h and that its absorption is poor in the absence of a delivery system (63). In addition, mRNA is a naturally unstable molecule that is easily broken down by endonucleases, 3' exonucleases and 5' exonucleases (64). One of the most challenging applications of mRNA vaccines for cancer is getting the mRNA into a sufficient number of cells with sufficiently high translation levels. This requires highly targeted and effective mRNA delivery mechanisms (65). Thus far, the identified delivery vectors for cancer mRNA vaccines include LNPs, polymeric vectors, peptide vectors, DC vectors, extracellular vesicle vectors and hybrid vectors.
One of the most well-developed, promising and widely used mRNA non-viral delivery methods is LNP. It is made up of four components that function together to encase and shield the delicate mRNA core: Cholesterol, helper phospholipids, cationic or ionizable lipids and polyethylene glycol (PEG) ylated lipids (66,67). Cholesterol is used as a stabilizer to increase LNP stability, ionizable cationic lipids can promote the autonomous aggregation of mRNAs to form a particle of ~100 nm and release mRNAs in the cytoplasm through ionization, and PEG can extend the half-life of the LNP complex (34,68). Natural phospholipids support the nanoparticles to form a lipid bilayer structure. Currently, there are LNP-mRNA cancer vaccines in clinical trials, e.g., mRNA-4157.
Purification of mRNA vaccines
The product of in vitro transcription of mRNA is primarily a combination of targeted mRNA, non-targeted mRNA, nucleotides, oligonucleotides and proteins. Chromatographic methods are often used to separate the target mRNA from other mRNA impurities in this system, while precipitation and extraction techniques are utilized to eliminate common impurities from the mRNA (69,70). Type I IFN production may be enhanced by dsRNA, a simulant of RNA virus genome replication intermediates. Consequently, mRNA translation efficiency may be increased and the type I IFN immune response to mRNA vaccines is effectively reduced by purifying an mRNA in vitro synthetic product. During IVT, dsRNA species may be decreased by either generating RNA at a higher temperature or by lowering the Mg2+ content (71). The purification characteristics of dT 25-conjugated oligonucleotide affinity support resin (dT25-OAS) were examined by Engel et al (72); due to its very high binding capacity, dT25-OAS is a desirable substitute for mRNA purification on a wide scale.
Molecular stabilization of mRNA vaccines
Increasing protein expression often involves techniques such as modifying sequences and/or the structure to improve mRNA stability (increase the half-life) and translation. Methods include extension of the poly(A) tail, alterations of the 5' cap, engineering untranslated region (UTR) and open reading frame (ORF) sequence patterns and altering the specific nucleotide sequence (30,73,74). These changes result in the production of considerable quantities of the protein for an extended period of time, ranging from a few minutes to >1 week (75-77). PERSIST-seq, a platform based on RNA sequencing, was created recently to comprehensively characterize the stability of intracellular mRNA of various mRNA libraries. This platform has enabled and continues to enable computational trials for mRNA drug enhancement (78).
5' cap modification
The 5' cap is a protective structure that may trigger mRNA translation, prevent exonuclease cleavage and control pre-mRNA splicing and nuclear export, amongst other functions (79,80). The innate immune system uses the 5' cap to distinguish endogenous RNA from exogenous RNA (81). Regarding in vitro mRNA capping, there are two standard methods. First, mRNA capping and in vitro transcription may be accomplished by including the normal cap analog, m7GpppG structure, into the mRNA transcription system. Second, after the first in vitro transcription, methyltransferases methylate Cap 0 to Cap 1 for mRNA capping (82).
The most commonly used capping technique for mRNA transcription in vitro is capping using cap analogs. Several cap analogs have now been developed. The anti-reverse cap analogs (ARCAs), which are altered inside the ribose moiety of the m7G, are the most often reported cap analogs (83). To enhance the quality of mRNA, additional alterations to the ARCA structure have been developed recently. By increasing the affinity of mRNA for eIF4E, phosphorus modification based on ARCA, for example, may increase mRNA stability by reducing its sensitivity to decapping enzymes and improving translation efficiency (84-87). The insertion of a novel chemically altered cap analog into ARCA increases the mRNA's half-life by preventing it from being decapped by mRNA decapping enzyme 2 (30). In 2018, 'CleanCap', a co-transcriptional capping technique, was created as an additional cap analog. Compared to first-generation cap analogs (such as mCap and ARCA), which have cap 0 structures at lower efficiencies and reaction yields, this was far more efficient (88).
Optimization of 5' and 3'-UTRs
The upstream (5'-UTR) and downstream (3'-UTR) domains of the mRNA coding region include the noncoding portion of the mRNA sequence or UTR. UTRs have several functions, including the control of mRNA stability, subcellular localization, translational efficiency and mRNA export from the nucleus. The mRNA expression levels in vivo are often increased by UTR optimization, hence UTR is a key part of cancer mRNA vaccine design (89-92).
Codon optimization of the ORF
The ORF region is the coding region of mRNA, hence its translation rate is unquestionably important. Therefore, the total translation efficiency of mRNA may be maximized by selecting the right codons in this region (93). To accomplish the goal of optimizing the sequence, either choosing the optimal codon pair that is common to the highly expressed protein or maintaining the same percentage of each codon that naturally exists in the highly expressed protein in the target cell are the methods used. Furthermore, to speed up translation, less common codons in the ORF are often replaced by codons with increased transfer RNA (tRNA) abundance (94). In such an instance, it is possible to ensure sufficient levels of tRNA during the production of foreign mRNA and/or the increased translation of highly expressed genes utilizing the host's codons (95). To control the translation elongation rate, the ORF's guanine (G) and cytosine (C) content may be optimized. Another codon optimization technique that has a direct correlation to a higher GC content is uridine depletion. RIG-I can identify areas rich in uridines, and when it is activated, protein expression may be completely stopped (49).
Poly(A) tail modification
At the 3' end of an mRNA construct is a polyadenylation region known as the poly(A) tail. It is important for both the enzymatic stability of mRNA and its translation. To control translational efficiency, it specifically binds to several polyadenosyl binding proteins and cooperates with the 7-methylguanosine cap (m7Gppp) on the 5'-end of mRNA sequences (96). According to early research, the poly(A) tail is crucial in controlling the stability and translation efficiency of mRNA. It has been discovered that longer tails result in increased protein expression levels in a variety of cell types. Shorter poly(A) sequences, on the other hand, may encourage this closed-loop shape for effective translation, according to research by Lima et al (97). In conclusion, to maximize the translation efficiency of mRNA, the length of the poly(A) sequence should be modified for various cell types.
Modified nucleotides
Certain mRNA nucleotides undergo post-transcriptional modifications during mRNA maturation. IVT mRNA may be synthesized using these naturally occurring modified nucleotides, such as 5-methylcytidine and pseudouridine. IVT mRNA exhibits better translation efficiency and stability when it possesses modified nucleosides such as pseudouridine. This is hypothesized to be due to the fact that nucleosides prevent TLR, RIG-I and The RNA-dependent protein kinase R (PKR) from being activated, making IVT mRNA undetectable to cytoplasmic TLRs such as TLR3, TLR7 and TLR8, as well as RIG-I and PKR (98). However, it has been suggested that unmodified mRNA vaccines can assist the immune system in the recognition of tumor cells, in which case modifications of cancer mRNA vaccines are not required.
Adjuvants
Adjuvants are substances, either natural or artificial, that the immune system can easily detect and use to augment the intended immunological response (99). The injection of its matching adjuvant may improve the immune response to antigens, even if mRNA vaccines themselves have a self-adjuvant effect. Adjuvant addition to mRNAs is a popular topic of research currently.
An mRNA adjuvant called TriMix consists of three immune-modulatory molecules: CD70, CD40 ligand and active TLR-4. Patients with stage III or IV melanoma were administered TriMix mRNA along with additional tumor-antigen mRNAs, and this resulted in long-lasting clinical relief and an augmented increased immune response (100,101). The adjuvant pulsed mRNA vaccine NP technique for c16-r848 was recently established. This technique significantly improved the amplification of ova-specific CD8+, which allowed the activated T lymphocytes to penetrate the tumor bed in vivo, something that was not observed with the mRNA vaccine NP without adjuvant (100). The RNActive (curevac Ag) vaccination platform and RNAdjuvant are other popular adjuvants. Furthermore, certain mRNA delivery vectors, such as protamine and cationic lipids, may enhance the effect of adjuvants. Pam2Cys, a synthetic neutral fatty amino acid that can signal through the TLR2/6 pathway, has been shown to trigger both humoral and cellular applicative immune responses, which makes it a promising candidate adjuvant (102). A recent study integrated Pam2Cys into mRNA-LNP to improve the efficacy of mRNA vaccines. In prophylactic and therapeutic tumor models using this vaccine, CD4+ and CD8+ T-cell-dependent anti-tumor responses were significantly enhanced, while memory anti-tumor responses were also established. Thus, Pam2Cys is of notable value in the development of future cancer mRNA vaccines (103).
4. Applications of mRNA vaccines in infectious diseases
mRNA vaccines are primarily being studied for the treatment of cancer and infectious disorders. At present, there are ~140 clinical studies assessing the use of mRNA vaccines to treat various illnesses (39). Self-amplifying or replicon RNA vaccines and non-replicating mRNA vaccines are the two primary types of RNA vaccines that have been assessed. Replicons are self-amplifying mRNAs that are produced from RNA viruses that have had their structural viral proteins replaced with mRNA-encoding RNA polymerases and antigens. Thus, these mRNAs enhance immunogenicity and extend protein expression, thereby improving efficiency (104,105).
Self-amplifying mRNAs have two ORFs, one encoding the targeted antigen sequence and the other encoding the viral replication mechanism, in contrast to the traditional non-replicating IVT mRNA of 'mature' eukaryotic mRNA. This allows for long-lasting RNA amplification in cells (49). The delivery mechanism of non-replicating mRNA vaccines may be further identified, since it can be administered directly into a range of anatomical areas or by ex vivo loading of DCs (34). While there are now several opportunities for the therapeutic use of self-amplifying mRNAs in the prevention of infectious illnesses, its use as a cancer vaccine is mostly restricted to preclinical research and has only seen a small number of clinical studies conducted (13,49).
Diseases caused by viral infection
Nucleic acid-based vaccines induce humoral and cytotoxic T-cell responses by imitating a viral infection and expressing vaccine antigens in situ during immunization (106). This benefit is essential for eliminating infections or intracellular pathogens, where strong humoral and cellular immune responses are needed to provide effective protection (Fig. 5).
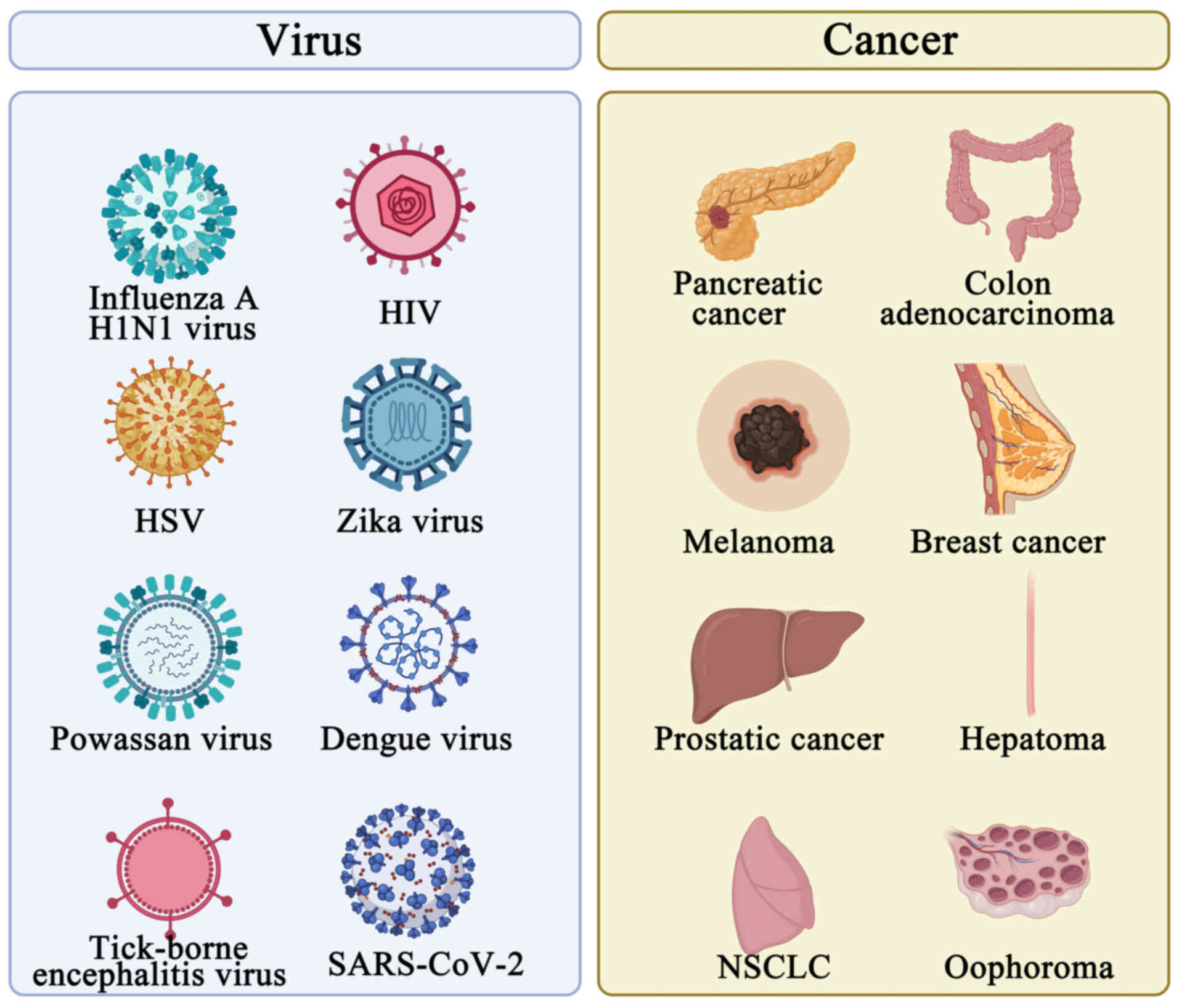
Influenza-causing virus
The most effective method to avoid influenza is vaccination (107). Compared to traditional vaccines, mRNA vaccines that encode the conserved regions of the influenza virus's effector protein(s) may induce the production of specific antibodies, improving prevention and ideally treatment outcomes (107). Furthermore, mRNA vaccines are simpler to develop for novel influenza viruses due to a speedier development process (108). This suggests that mRNA vaccines may serve as quick and adaptable tools for managing influenza, both seasonal and pandemic.
Human immunodeficiency virus (HIV)
In the first human clinical trial with naked mRNA (ihivarna) in combination with a novel HIV immunogen sequence (HTI immunogen) using a DC activation approach (trimix: CD40 ligand (CD40L)+ CD70+ encode constitutively active TLR4 (CATLR4) RNA), and vaccination, the vaccine was safe and well tolerated. It rapidly increased HIV-1 DNA and RNA in peripheral CD4 T cells and triggered a modest HIV-specific T-cell response (109). The activating adjuvant trimix and 16 conserved segments (GAG, pol, Vif and Nef) from the HIV-1 structural protein were used to make up the HTI trimix. It encoded a potent activation signal and an efficient HIV recombinant antigen, making it a promising novel vaccine candidate for mRNA-based therapy of HIV-1. According to preclinical findings, it may successfully stimulate T-cells, the release of mature DCs and the release of antiviral cytokines (particularly IFN-γ). Phase I and phase IIa clinical studies of HTI-TriMix were completed by the end of 2019. HIV-1-positive individuals in a phase IIa study were administered three vaccines at weeks 0-2 and 4, as determined by ultrasonography guidance using an inguinal lymph node (109,110).
Herpes simplex virus (HSV)
A trivalent vaccination that targets the glycoproteins C, D and E found on HSV-2 viral particles was developed recently. After an HSV-2 assault, the vaccination may shield animals against genital infection and subsequent viral shedding. In addition, vaccination generated cross-reactive antibodies, which neutralized HSV-1 and provided defense against HSV-1 infection. As a result, this vaccine offers a strong defense against genital HSV-1 and HSV-2 infections and is a suitable candidate vaccine for human testing (111) (Fig. 5).
Flaviviruses
Numerous flaviviruses, such as Zika virus, Powassan virus, dengue virus and tick-borne encephalitis virus, have been treated and infections prevented using mRNA vaccines (112-116). The use of mRNA vaccines against the Zika virus has garnered significant interest. To assess immunogenicity and protection in mice, Richner et al (115) created an LNP-encapsulated modified mRNA vaccine containing wild-type or mutant ZIKV structural genes. In a phase I/II human clinical study, a comparable vaccine called mRNA-1893 resulted in >90% seroconversion after a prime-boost immunization administered at dosages of 10 or 30 µg (113) (Fig. 5).
COVID-19
The use of mRNA vaccines against COVID-19 has garnered notable interest. To date, several vaccines have been produced, including mRNA-1273, BNT162, ARCT-021 and the cvncov vaccine (cv07050101). The near-identical 94 to 95% vaccination efficacies of the mRNA-1273 and BNT162b2 COVID-19 vaccines, as well as their rapid development and testing period of a year, are remarkable achievements in science and medicine (117).
Among the mRNA vaccines under development, only the mRNA vaccine for COVID-19 is currently on the market. Based on published clinical data, the COVID-19 mRNA vaccine provides a high level of protection and safety. With mRNA vaccination, recipients have significantly higher levels of neutralizing antibodies than those who have recovered from COVID-19 and who were not vaccinated and exhibit an enhanced immune response and memory. The mRNA COVID-19 vaccine provides 95% overall protection against the virus in individuals aged ≥16 years and can effectively induce a response to 20+ variants of the virus. Clinically significant safety concerns have not been identified in the majority of patients receiving the mRNA COVID-19 vaccine. Myocarditis after neocoronary mRNA vaccination has been observed in a small percentage of those vaccinated, with the highest incidence in younger men after a second dose of the vaccine (118).
Diseases caused by bacterial infection
Attempts to develop mRNA vaccines against bacterial and parasitic antigen species are limited, many of which are still in the preclinical testing stage.
Maruggi et al (119) examined the effectiveness and immunogenicity of self-amplifying mRNA vaccines that express group A streptococcal antigen (GAS) and GBS. Self-amplifying mRNA vectors were able to effectively produce two prototype bacterial antigens: The GBS pilus 2a backbone protein and the double-mutated GAS streptolysin-O. In mice infected with GAS and GBS, the antibody responses produced by self-amplifying mRNA vaccines were able to consistently provide protection. Jawalagatti et al (120) developed a novel strategy for creating mRNA vaccines to be taken orally.
5. Applications of mRNA vaccines in various tumors
Approximately 25 years ago, the viability of an mRNA-based cancer vaccine was initially shown (121). Since then, the use of mRNA vaccines in cancer treatment has been investigated in several preclinical and clinical investigations. A small number of mRNA cancer vaccine clinical studies have been conducted so far (122). The majority of trials have focused on acute myeloid leukemia (AML) (123), multiple myeloma (124), mesothelioma (125), glioblastoma (126), malignant glioma (127) and renal cell carcinoma (128), as well as mRNA vaccines for pancreatic cancer (129), melanoma (130), BCa (131), non-small cell lung cancer (132), prostate cancer (133), ovarian cancer (134) and colorectal cancer (135) and are still in their early stages (I and II) (136-140) (Fig. 5). Moderna and Merck Sharp & Dohme announced that the first combination therapy of an mRNA personalized cancer vaccine (mRNA-4157) and an anti-programmed cell death 1 (PD-1) monoclonal antibody (Keytruda) advanced to phase III clinical trials in July 2023 (14). This was the first mRNA cancer vaccine to enter phase III clinical trials. Table I summarizes the clinical trials of mRNA vaccines for the treatment of cancer.
Pancreatic cancer
Growing data suggest that the intra- and intertumoral heterogeneity of pancreatic cancer, particularly in relation to the immune microenvironment and genetic alterations, is associated with resistance to treatment (141-143). Traditional cancer treatment approaches often result in the acquisition of resistance, underscoring the need for targeted, individualized care (140). In patients with pancreatic ductal carcinoma, a recent phase I clinical study using the adjuvant tailored mRNA neoantigen vaccine autogene cevumeran, the vaccination stimulated T-cell activity, which may have been associated with a delayed recurrence of the illness. In the clinical trial, 16 of 19 patients were treated with cevumeran after treatment with Atezolizumab; 8 of the 16 patients who received the mRNA vaccine produced T-cell translocations that sustained the production of IFNγ, demonstrating the immune persistence of the mRNA vaccine. The median relapse-free survival (RFS) of subjects who responded to the vaccine had not been reached at the median follow-up time of 18 months. The RFS was significantly higher in responders than in non-responders. Therefore, it can be concluded that Atezolizumab (an anti-PD-L1 monoclonal antibody) in combination with the mRNA vaccine induced extensive T-cell activity and delayed the recurrence of pancreatic cancer (129). In patients with pancreatic duct adenocarcinoma (PDAC) undergoing surgical resection and at risk of delayed recurrence, Kang et al (144) demonstrated that the personalized mRNA neoantigen vaccine, autogene cevumeran (BNT122), induced significant T-cell activity when combined with Atezolizumab and mFOLFIRINOX, a standard adjuvant chemotherapeutic regimen following surgery for PDAC.
Colon cancer
mRNA vaccines against colon adenocarcinoma are limited. A significant difficulty is sorting through the several vaccine candidates to identify possible mRNA vaccines that may be suitable for a specific type of cancer (145). Thrombospondin 2, follistatin like 3, troponin T1, biglycan, collagen triple helix repeat containing 1 and NADPH oxidase 4 are among the targeted antigens that Liu et al (139) identified as potential mRNA vaccine candidates. The study also described aberrant gene expression patterns and the mutational landscape of CRC. The results may assist in the determination of patients with CRC who are suitable candidates for immunization and provide a theoretical foundation for the creation of mRNA cancer vaccines.
Melanoma
To develop mRNA vaccines and identify appropriate vaccine populations, Ping et al (146) analyzed potential tumor antigens in melanoma. Their analysis revealed that protein tyrosine phosphatase receptor type C, sialic acid binding Ig like lectin 11 (SIGLEC11), cyclic-oligoadenylate-activated single-stranded ribonuclease and single-stranded deoxyribonuclease 1, leukocyte immunoglobulin-like receptor subfamily B member 1 and ADAM like decysin 1 may be antigenic targets for mRNA vaccines for melanoma, laying the groundwork for the development of mRNA vaccines in melanoma (146). According to the findings of Sittplangkoon et al (147), unaltered mRNA vaccines in melanoma models may result in the production of IFN-I or trigger downstream signaling cascade responses, both of which are essential for eliciting strong anti-tumor T-cell responses that regulate tumor development and metastasis. The experimental vaccine mRNA-4157/V940 in combination with pembrolizumab may be used as an adjuvant therapy for the treatment of high-risk melanoma, according to the results of a recent clinical study. Compared with patients treated with a PD-1 inhibitor alone, individuals receiving this combination treatment had a noticeably decreased risk of recurrence after surgery (130).
BCa
Li et al (148) identified groups of patients with BCa that may be suitable candidates for mRNA vaccination as well as putative BCa-associated antigens for the creation of anti-BCa mRNA vaccines. A total of three tumor-associated antigens were identified: CD74, interferon regulatory factor 1 and proteasome activator subunit 2. These antigens were frequently mutated, amplified or upregulated, and they were associated with immune-cell infiltration and prognosis. The tumor microenvironment (TME) in patients with class B immune subgroups may respond well to mRNA vaccines (148).
Patients in the prostate adenocarcinoma (PRAD) immune subtypes 2 and 3 groups were more likely to benefit from immunization, and prospective antigens for the PRAD mRNA vaccine, including Kelch-like family member 17, carnitine palmitoyltransferase 1B, IQ motif containing GTPase activating protein 3, Lck interacting transmembrane adaptor 1, YjeF N-terminal domain containing 3, coiled-coil domain containing 180, MutS homolog 5 and cadherin EGF LAG seven-pass G-type receptor 3 (133).
Hepatocellular carcinoma (HCC)
Peking Union Medical College Hospital is conducting the first human clinical trial of the tumor antigen mRNA therapeutic vaccine ABOR2014 (IPM511). The vaccine encodes nearly 20 HCC antigens that are frequently upregulated in patients with HCC. The study aims to evaluate the safety, tolerability and preliminary efficacy of IPM511 alone and in combination with PD-1 inhibitor therapy in patients with advanced HCC with disease progression after first-line standard therapy. The study is currently ongoing and no results have been published, with the principal investigator stating that IPM511 has been shown to demonstrate safety and clinical accessibility in the first patient treated using this approach
Esophageal cancer
A study of a personalized mRNA cancer vaccine combined with a PD-1 inhibitor for the treatment of advanced esophageal squamous cell carcinoma was recently reported (149). The patient enrolled in the study was first screened for neoantigens using whole-transcriptome sequencing of tissue samples from the lesion. A personalized mRNA cancer vaccine was developed that was tailored to these neoantigens. Subsequently, the patient was administered the mRNA cancer vaccine in combination with a PD-1 inhibitor and the patient obtained partial remission; the safety of the treatment regimen was within manageable limits. Thus, mRNA vaccines may be an effective therapeutic strategy for patients with advanced esophageal cancer.
6. Functional implementation of mRNA vaccines in tumor treatment
Immunotherapy of mRNA vaccines
Cancer may now be effectively treated with immunotherapy, and given the positive early clinical outcomes, the FDA has authorized an increasing number of immunotherapies. The foundation of cancer immunotherapy is the immune system's capacity to identify and eliminate cancer cells (150). With the advent of several classes of medicines targeted at augmenting immune responses against malignancies, this area of research has rapidly expanded in recent years. These consist of various vaccination approaches, adoptive T-cell treatments, immune checkpoint inhibitors and cytokines (151-153).
Tumor antigens
Tumor-associated antigens (TAAs) and tumor-specific antigens (TSAs) are two categories of antigens that are encoded by mRNAs and have traditionally been the target of immunotherapy. Given immunologic memory, vaccines that specifically target TAAs or TSAs may target and kill cancerous cells that exhibit upregulated expression of antigens and provide a long-lasting therapeutic response (49).
TAAs
TAA is not unique to tumor cells. While they may also be found in healthy tissues and cells, particularly in embryonic tissues, the expression of these antigens is markedly elevated in tumors. TAAs in immunotherapy are potential targets as shown by Conry et al (154) who developed the first mRNA cancer vaccine; this vaccine demonstrated that mice immunized with mRNA coding for carcinoembryonic antigen (CEA) showed an anti-CEA antibody response when challenged with CEA expressing tumor cells. Significant attention has been given to TAA research since this initial discovery, with positive clinical study outcomes. Three tumor antigens, including CD247, Fc gamma receptor Ia and transcription domain associated protein, were identified by Huang et al (155) in cholangiocarcinoma. These antigens are associated with an improved prognosis and the infiltration of antigen-presenting cells. These antigens may serve as potential candidates for mRNA vaccines against cholangiocarcinoma (128,155). TAA immunization methods do have certain drawbacks, however. The TAAs are often not found in malignant cells. In addition, as not all discovered TAAs induce an antitumor immune response, selecting TAAs may be challenging and vaccine development is expensive. There is a possibility that the tumor itself may downregulate the TAA, allowing for escape when focusing on a particular antigen (98). Finally, TAAs may also be found in normal tissues, and vaccines against them may cause peripheral and central tolerance responses, and this may result in poor vaccination efficacy or autoimmunity against normal tissues (156).
TSAs
TSAs, also known as neoantigens, are antigens unique to tumor cells from somatic random changes and are thus absent from normal tissues and cells. Tumor-specific mutations are appealing targets for cancer immunotherapeutics and these mutations may result in novel antigens, allowing for the development of customized vaccines, as the mutated antigens are not expressed in healthy tissues and may thus be identified by T-cells (157,158). The following characteristics of neoantigen-based vaccinations highlight their benefits over TAA-based vaccines. Since neoantigens are only produced by tumor cells, they can only trigger T-cell responses specific to tumors, limiting harm to non-malignant tissues. Neoantigens offer the potential to elicit immunological responses against tumors by evading T-cell central tolerance of self-epitopes. Furthermore, there may be long-term protection against disease recurrence due to the capacity of these vaccine-boosted neoantigen-specific T-cell responses to endure and provide immunological memory beyond therapy (159). To immunize patients against metastatic gastric cancer, Cafri et al (160) recently linked the verified and characterized novel antigens and predicted new epitopes and driver gene alterations into a single mRNA construct. The developed vaccine proved safe and produced a T-cell response specific to the expected new epitopes that had not been discovered before immunization. The use of neoantigens still faces several challenges despite the advancements made; these obstacles stem from both biological and technical factors, including polymorphism of putative antigens and HLA molecules, gaps in our understanding of HLA binding motifs for less common HLA alleles and the heterogeneity of tumors (161-164).
Delivery methods of mRNA vaccines for tumor antigen
mRNA-transfected DC vaccines
DCs may be transfected with the mRNA expressing tumor antigen and total tumor mRNA, which can subsequently be delivered to the host and initiate an immune response. Subcutaneous application of DCs transfected with TAA mRNA or total mRNA to tumor-bearing mice has been shown to induce T-cell immunity and inhibit the growth of established tumors as early as the last century, which also supports the feasibility of these two distinct transfection methods (61). Patients receiving docetaxel for metastatic castration-resistant prostate cancer showed no adverse effects when Kongsted et al (165) transfected DCs expressing numerous TAAs; ~50% of the research participants exhibited an immune response. A phase 1 clinical trial including patients with lung cancer is now assessing MIDRIXNEO, a customized mRNA-loaded dendritic cell vaccine that targets tumor neoantigens (166).
Direct injection
There are several methods to directly inject the mRNA encoding the tumor antigen into a host. Local cells, such as APCs, take up the mRNA and translocate it to the cytoplasm where it is translated. One of the most common methods of delivering tumor antigens is via intranodal injection. Phase I research on 29 patients with advanced melanoma started in 2012; unpublished data from this trial showed that intranodal mRNA injection is safe and viable (NCT01684241). mRNA encoding tyrosinase, premelanosome protein, MAGE family member A3 (MAGE-A3), MAGE-C2 and melanoma antigen preferentially expressed in tumors together with TriMix mRNA were well tolerated when injected intravenously. In another phase I clinical study, intranodal delivery of a neoantigen-specific mRNA vaccine, consisting of 20 mutations specific to the patient, was studied in patients with stage III and IV melanoma (NCT02035956), which demonstrated that individual mutations can be exploited (167).
Administration via the skin
Administration of tumor antigens expressed by an mRNA may be achievable via the skin. Mice were injected with IVT mRNA encoding the enhanced green fluorescence protein and the melanocyte autoantigen TRP2 by blasting their skin with a gene gun. This successfully elicited cellular immunity specific to antigens, mediating a protective effect against B16 lung metastases and inducing vitiligo-like hair decolorization (168). Protamine-condensed naked mRNA encoding six melanoma-associated antigens was safely and successfully injected intradermally into two of the four evaluable patients in a phase I/II clinical trial involving 21 patients with metastatic melanoma. In addition, 1 of the 7 patients with measurable disease showed a complete response to the vaccine (169).
Intranasal administration
The mucosal route, which encodes a tumor antigen via mRNA, may efficiently elicit both a systemic immune response and localized mucosal immunity. Nasal-associated lymphoid tissue, a favorable location for antigen internalization to promote protective responses against cancer cells, may be reached via intranasal delivery. In an invasive Lewis lung cancer model, Mai et al (170) immunized mice intraperitoneally with a cationic liposome/protamine complex carrying mRNA expressing cytokeratin 19, and this slowed down the tumor's development and elicited a robust cellular immune response.
Intravenous administration
The melanoma fixvac (bnt111) intravenous liposome RNA (rna-lpx) vaccine, which encodes four non-mutated tumor-related antigens, was investigated by Sahin et al (171). The rna-lpx vaccination was a successful method of immunotherapy; robust CD4+ and CD8+ T-cell immunity was achieved against the vaccine antigen, which was accompanied by a favorable clinical response. In certain responders, the antigen-specific cytotoxic T-cell response was durable and to the degree of often documented adoptive T-cell treatment.
mRNA-encoded immunomodulators
Tumor immunotherapy, also known as active nonspecific immunotherapy, was one of the first fields of medicine to use immune system modulators. Target cell transfection in vitro, or intravenous or intratumoral delivery, are the primary methods for producing immunomodulators via mRNA in vivo (121). The immune system's cells can communicate across short distances due to cytokines, which are important regulators of both innate and adaptive immunity. To stimulate the immune system of patients with cancer, cytokine therapy has been a significant therapeutic approach and is a major topic of clinical cancer research at present (172). The goal of mRNA-encoded cytokine-based immunotherapy is to increase the quantity of cytokines in the TME with the least amount of toxicity and systemic exposure possible from the delivery of recombinant proteins (173).
Given its strong proinflammatory properties, IL-12 is a promising option for cancer immunotherapy. Type 1 T-helper cell (Th1) differentiation, the acquisition of cytotoxic activities by CD8+ cells, and the activation of IFN-γ production, all of which improve phagocytic functions and local inflammation, are all induced by IL-12 signaling (174). T-cells transfected with mRNA producing a single-chain IL-12 (scIL-12) made up of the p35 and p40 subunits were the subject of one investigation. In syngeneic and xenograft mouse models, intratumoral injection of scIL-12-expressing T-cells led to total rejection of both injected and remote tumor lesions. The anti-tumor efficaciousness of T-cells was further enhanced by co-electroporation with 4-1BB ligand (4-1BBL) mRNA (175). It has been shown that novel intratumoral IL-12 mRNA treatment can stimulate Th1 TME transformation and powerful antitumor immunity (176). Additionally, in mouse tumor models resistant to checkpoint inhibitors, it was shown that the combination of immune checkpoint inhibitors with IL-12 mRNA increased the anticancer response, enhancing overall survival (OS) and tumor regression (176).
Multiple mRNAs encoding various cytokines have been validated for their immunotherapeutic effects. The mRNA encoding four cytokines, IL-12, IFN-α, IL-15 sushi, and granulocyte-macrophage colony-stimulating factor, was examined by Hotz et al (177) after intratumoral administration. These cytokines may promote the development of immunological memory and have potent anticancer action. Hewitt et al (178) showed that in a variety of TMEs, direct intratumoral administration of mRNAs encoding these cytokines generated strong anticancer responses.
Tumor immunotherapy also involves mRNA-encoded stimulatory ligands and receptors, in addition to cytokines. The immune system receives inflammatory signals from stimulatory ligands and receptors, which may be used in cancer immunotherapy. By temporarily supplying cells with stimulatory receptors, mRNA may be used to temporarily activate potent inflammatory signals. While these immunostimulants are not regarded as cancer vaccines, they are often administered in conjunction with other immunotherapeutic treatments, such as checkpoint blockade modulators, to enhance the humoral and cellular responses. Several studies showed that the immunostimulatory activity of DCs was significantly increased upon electroporation with mRNAs encoding co-stimulatory molecules, including CD83, TNF receptor superfamily member 4 (also known as OX40) and 4-1BBL. Trimix mRNA, an mRNA-based adjuvant that was created in 2016, is made up of three mRNA molecules that encode CATLR4, activating stimulator CD40L, and costimulatory molecule CD70. Intramuscular injection of tethered IVT mRNA-TLR7 agonists boosted antigen-specific cell-mediated and humoral responses in vivo, as shown by Loomis et al (179). Both alone (TriMix mRNA plus TAA mRNA, or autologous monocyte-derived mRNA co-electroporated dendritic cells with mRNA encoding CD40 ligand, CD70 and a constitutively activated TLR4, which is referred as TriMixDC-MEL) and in combination with ipilimumab checkpoint inhibitor, a monoclonal antibody that blocks CTLA, the products were able to elicit a potent immune response, which in turn led to a promising clinical response and prolonged disease-free survival rates (NCT01676779 and NCT01302496) (101,180). These phase II studies were conducted for the treatment of patients with stage III/IV melanoma.
mRNA-encoded antibodies
Since the first antibody was licensed for use in cancer therapy, the field of oncology has developed antibody-based therapeutics at a progressively faster pace. As a result, there are now several approved antibodies and additional candidates that are undergoing clinical review (181). The ability of mRNA to produce various antibodies and antibody types in vitro was shown by Thran et al (182). furthermore, the mRNA that encodes tumor antibodies may elicit robust anti-tumor immunity.
The following methods may be used to categorize the involvement of antibodies in tumor treatment: i) Antibodies that, after binding to the target receptor, may facilitate direct death of tumor cells by inducing apoptosis signals or removing vital growth signals. ii) Complement dependent cytotoxicity (CDC) or antibody-dependent cell-mediated cytotoxicity (ADCC) are induced when immune-mediated cell death is activated by attaching to antigens specific to cancer cells. iii) Blocking the immunological checkpoint, which involves preventing the antibody of PD1 or CTLA4, reactivating the T-cell response specific to the antigen against cancer cells, and triggering immune-mediated cell death. iv) Directly engaging T-cells with cancer cells to initiate immune-mediated cell death (183-188).
Monoclonal antibodies (mAb), mAb fragments and engineering variations (such as diabodies, triabodies, minibodies and single-domain antibodies) are the categories of antibodies that are involved in tumor immunotherapy. Both monoclonal and bispecific antibodies have been investigated and used extensively (181).
The first mAb licensed for cancer therapy, rituximab, targets CD20 and induces CDC and, to a lesser degree, ADCC. It is used to treat chronic lymphocytic leukemia and non-Hodgkin's lymphoma (189). A technique that has been beneficial in a preclinical B-cell lymphoma model is the encoding of this mAb in mRNA (190). Cancer cells starve and eventually die as a result of trastuzumab's inhibition of kinase activity, downstream signaling and human EGFR 2 (HER2)/Erb-B2 receptor tyrosine kinase 2 receptor dimerization (186). Direct tumor cell death may also be induced by antagonistic antibodies binding to apoptosis-inducing receptors on cancer cells, such as TNF-related apoptosis-inducing ligand receptors and antibody-drug conjugates that deliver toxic payloads, particularly to cancer cells (184,188). When intrathecally administered using a liver-targeting LNP formulation, trastuzumab encoded by mRNA was retrieved from mouse serum and exhibited ADCC. In a HER2/neu-positive BCa xenograft model, this approach increased survival (191). The translation of anti-programmed cell death ligand 1 (PD-L1) mAb in transfected cells and the infiltration of CD8+ T-cells into tumors may be facilitated by injecting antagonistic anti-PD-L1 mAb expressing self-amplified RNA into mouse tumors (192). Despite the several positive instances of mAbs in cancer, poor tissue penetration limits the ability of the complete antibodies to distribute uniformly across the tumor mass. As a result, efforts have been made to modify the structure of antibodies, resulting in the creation of novel antibody fragments with improved tissue penetration (173). The discovery of antibody fragments has considerably aided in the study of bispecific antibodies (BsAbs) and these have been employed as building blocks in the creation of these antibodies. The world's first antibody to bind two antigens simultaneously was created in the 1960s by Nisonoff et al (193) by combining the antigen binding fragments from two rabbit polyclonal antibody sera using a moderate reoxidation approach (194,195).
One possible benefit of BsAbs over regular mAbs is their ability to target two distinct sites simultaneously and, to a certain degree, circumvent tumor drug resistance. However, since BsAbs have short serum half-lives (just a few hours) they must be constantly administered to the patient using an infusion pump (196). One possible method to overcome this restriction is to employ mRNA to directly generate therapeutic antibodies in patients.
A recent study highlighted a successful bispecific targeting method; a clinically authorized LNP encapsulating mRNA expressing C-C motif chemokine ligand 2 (CCL2) and CCL5 (bisccl2/5I), which can bind and neutralize bispecifically. This markedly promoted TAM polarization towards the anti-tumor M1 phenotype and lowered immunosuppression in the TME. BisCCL2/5i in conjunction with PD-1L inhibitor has been shown to result in long-term survival in a mouse model of pancreatic, colorectal and liver metastases from primary liver cancer (193). The use of mRNA encoding a BsAb with ablation Fc immunological impact function was described by Wu et al (197). The antibody, also known as XA-1, targets human PD-L1 and PD-1. In contrast to direct antibody therapy for cancer, this work demonstrated that treatment with XA-1 mRNA LNPs may efficiently produce endogenous therapeutic BsAbs via hepatocytes.
mRNA-encoding antigen receptors
By producing transgenic T-cell receptors (TCRs) or CARs, it has been possible to provide naïve T-cell tumor selectivity due to the transitory nature of mRNAs (173). mRNAs encoding CARs or TCRs have shown to be of value; generating transient expression of CARs or TCRs may prevent the development of cytokine release syndrome, a negative consequence of chronic T-cell activation (198-201). Considerable interest has been shown in CAR-T cells as the most promising cancer-adoptive immunotherapy approach.
With CAR-T treatment, immunological T-cells are extracted from patients, genetically altered in vitro, and endowed with a 'chimeric antigen receptor' (CAR) that identifies the antigen on the surface of cancerous cells. These altered cells are grown in a lab before being reinfused into the patient (202). Typically, extracellular TAA-specific antibody-binding domains are fused to intracellular T-cell-signaling regions to form chimeric receptors. Virtually every tumor-associated antigen may be targeted by CAR-T cells (203). There are several hazards associated with targeted non-tumor toxicity in CAR-T treatment. In vitro transcribed mRNA CAR-T cells are emerging as a safe therapeutic option, since they can avoid focused anti-tumor toxicity (204). The majority of studies use mRNA electroporation of CARs, as it is a practical and scalable method that achieves lymphocyte transfection rates of >90% without compromising the viability of the cell product (205-208). CAR expression is observed on the surface of T cells for ~7 days following mRNA electroporation, and CARs internalized upon target cell encounter are not restored (199,207-213). However, the IVT mRNA approach has drawbacks as well. These include poor tumor infiltration, manufacturing difficulties when a limited number of T-cells are available, the risk of side effects when repeated doses of CAR-T cells are injected and the short lifespan of mRNA-redirected T-cells, which results in the expression of encoded protein for a number of days (214). Most importantly, electroporation has various drawbacks that may significantly impact the caliber of the CAR-T cells generated (53). In fact, the integrity of the cell plasma membrane may be irreparably compromised by the application of pulsed electric fields. In the transfected cells that survive, decreased transgene expression, abnormal gene expression profiles and poor viability may be observed. In fact, it was demonstrated that when chemically modified 1mΨ mRNA and/or mRNA that was further purified to remove dsRNA was used, as opposed to their unmodified and unpurified counterparts, murine T-cells electroporated with CAR-encoding mRNA showed a markedly reduced upregulation of checkpoint molecules (PD-1 and lymphocyte activation gene 3). This gave the immunosilent mRNA-transfected T-cells a greater ability to kill, which persisted even after the cells' CAR expression was eliminated. The current results must be considered in the production of mRNA CAR-T cells to meet the objectives of the clinical trials, which are now conducting mRNA CAR studies for both hematologic and solid tumor malignancies (215).
Currently, mRNA CAR-T cells are receiving significant interest and showing promising outcomes in the study of tumor treatment. Treatment of hematological malignancies using mRNA CAR-T-cell therapy has seen favorable preclinical and clinical outcomes. Primary targets include CD19, CD37, CD33 and CD123. Preclinical research on mRNA CAR-T cell treatments for lymphoma and leukemia has been conducted (204). The approach for Hodgkin's lymphoma targeting CD19 and the approach for recurrent/refractory acute myeloid leukemia targeting CD123 are both being tested in clinical studies. These two studies demonstrate that mRNA CAR-T treatment is safe (204). A novel concept for mRNA CAR-T cell treatment was recently presented by Jetani et al (216), who identified siglec-6 as a new target of CAR-T cells in AML. Descartes-08, a novel CD8+ CAR-T cell product, was created by Lin et al (217) to treat multiple myeloma; preliminarily sustained responses and a strong treatment index were shown by the product. Mesothelioma, ovarian cancer, colorectal cancer, BCa and melanoma are among the solid malignancies for which mRNA CAR-T treatment has been investigated (204).
Combination therapy-immunotherapy and mRNA vaccines
mRNA therapy combined with another mode of treatment may show great promise in the treatment of tumors. The interaction between tumor antigens and immunomodulators is the primary focus of current research.
Early research has shown that TAA and IL-12 mRNA transfection into mature DC boosted TAA-specific CTLs, which may be exploited to trigger an immune response against tumors that is mediated by NK effector cells and CTLs (218). The safety and immunogenicity of cellular immunotherapy using trimixdc Mel were demonstrated by tests of mRNA encoding fusion proteins of a HLA-class II targeting signal (DC-LAMP), melanoma-associated antigen encoding the CD40 ligand, CATLR4 and CD70. At the study's IV dosage level, antitumor efficacy with long-lasting disease control was seen (219).
Sunitinib and rocapuldencel-T were recently employed in the phase 3 study for metastatic renal cell cancer. OS of patients receiving combined therapy was not improved by self-immunization via electroporation with amplified tumor RNA and CD40L RNA prepared from mature monocyte-derived DCs. However, the OS-related immune response was enhanced (220).
Protein therapy of mRNA vaccines
Methods of inducing cancer-cell death
Cancerous cells may be made to produce a deadly intracellular protein by employing mRNA therapies, which causes the cells to self-destruct (196). Van Hoecke et al (221) delivered mRNA encoding the mixed lineage kinase domain-like protein intrauterinally, which resulted in arrested tumor growth and induction of necrotic tumor-cell death.
Protein replacement therapy
The use of IVT mRNA in protein replacement therapy is predicated on the production of foreign proteins that can either activate or inhibit cellular pathways, as well as the supplementation of proteins that are absent or underexpressed. IVT mRNA injection has been used to target several proteins since its first preclinical assessment for protein replacement in 1992 (25).
Given their accessibility, the liver, lungs and heart are the primary targets of protein replacement treatments based on IVT mRNA. However, other tissues and organs, such as the skin, the back of the eye or the nasal cavity, have also been considered potential targets. Rare and hereditary disorders are the primary targets for this mode of treatment. The disease-causing down-regulated protein is linked to genetic abnormalities and encoded by the therapeutic IVT mRNA (222-227).
IVT mRNA as protein replacement therapy is being studied as a mode of cancer treatment (228). This is very difficult, however, as it requires recurrent administration, at times even systemic distribution, and targeted mRNA expression. In the realm of protein replacement, no clinical trials using IVT mRNA have been started yet, to the best of our knowledge (229).
Mutations of the oncogene tumor protein (TP53) are present in 96% of patients with high-grade serous ovarian cancer. p53 protein expressed by the TP53 gene recognizes and repairs DNA damage in cells and induces cell death when the damage is not repairable, thus avoiding the proliferation of aberrant cells to prevent the onset of cancer. When the TP53 gene is mutated, the mutant p53 protein that is expressed loses its oncogenic effect. In a recent study, a team synthesized mRNA encoding the correct p53 protein and loaded it into liposomes for delivery, thus allowing the mRNA to express functional p53 protein in cancer cells. After treatment with the above mRNA, patient-derived ovarian organoids began to shrink and die, and primary tumors and metastases in mouse models of ovarian cancer almost completely disappeared. These findings suggest that IVT-mRNA-based protein replacement therapy may reactivate oncogenic proteins in cancer cells and may thus serve as a valuable therapeutic option for the treatment of cancer (230).
Gene editing of mRNA vaccines
mRNA technologies expressing programmable nucleases, such as zinc-finger nucleases, transcription activator-like effector nucleases and clustered regularly interspaced short palindromic repeats (CRISPR)-CRISPR-associated endonuclease (Cas)9 systems (231), allow for gene editing as a potential therapeutic approach. By delivering site-specific alterations into a cell's genomes, such as correcting harmful mutations or introducing protective changes, these genome-engineering techniques allow for the replacement or modification of gene expression (232). CRISPR-Cas9 consists of Cas9 nucleic acid endonuclease and a single guide RNA (sgRNA) and is a widely used gene editing tool. CRISPR-Cas9 therapy can be used to permanently abrogate cancer cell survival genes, avoiding the need for repeat administration and thus improving treatment efficacy (233). Cancer cells or T-cells were transfected with Cas9 mRNA and sgRNA encapsulated in LNPs. The mRNA is translated in the cytoplasm to produce the Cas9 protein, which then complexes with sgRNA to form ribonucleoproteins with affinity for specific DNA sequences. Next, ribosomal complexes translocate to the nucleus to destroy tumor survival genes. When this happens in T-cells, PD-1 and endogenous TCR genes are knocked down to induce apoptosis in tumor cells (12). Ling et al (234) treated cervical cancer with Cas9 mRNA and gRNA targeting the E6 or E7 oncogenes. The results showed that this method not only effectively knocked down the oncogenes but also reversed the immunosuppressive microenvironment to achieve anti-tumor effects.
Regenerative medicine
The goal of regenerative medicine is to replace, regenerate or restore damaged or destroyed tissues, organs or cells by re-establishing or restoring their normal function (235). Proteins that regulate cellular biological activity, such as mitosis, migration and differentiation, and growth factors, cytokines and transcription factors, are essential to the regeneration process. An appealing substitute for traditional somatic cell reprogramming and transdifferentiation is the transfection of somatic cells with IVT mRNA expressing transcription factors (236). The primary goal of using IVT mRNA in regenerative medicine is to increase the ability of patients with type 1 diabetes to secrete insulin. The use of cell and mesenchymal stem cell engineering for tumor treatment has not received significant attention. It has been shown that stem cell exosomes with metastatic stem cell properties have been induced from stem cells using prostaglandin E2 receptor antagonists and used in cancer therapy and regenerative medicine. Stem cell exosomes can reduce tumor resistance (237). Whether it is possible to express stem cell exosomes with metastatic stem cell properties using mRNA vaccines for cancer treatment is a point for consideration.
7. Conclusion and future prospects
Patients with cancer have better odds of surviving thanks to the ongoing development of novel antitumor medications and therapies; however, a number of issues, including tumor drug resistance, dose-toxicity and other issues, keep posing new difficulties. In contrast to standard treatment methods, biotherapy for the majority of malignancies aims to prevent tumor growth and incidence by stimulating autoimmunity and obstructing critical signal transduction. There is some concern regarding the lack of long-term effects of mRNA-based technologies; researchers are attempting to challenge how illness is now treated using mRNA therapy.
mRNA vaccines have shown to be successful in several applications since Wolff et al (24) showed that mRNA transcribed in vitro in live cells may create proteins. As a promising novel platform, mRNA-based vaccines need to overcome the limitations of cold chain preservation and provide improved adaptability, efficacy, simplicity and scalability, as well as reduced costs. mRNA vaccines have shown benefits in infectious illnesses and cancer is the intended target of mRNA technology. Within the next 2-4 years, several active clinical studies using tailored cancer vaccines should be finished.
The use of combinatorial mRNA treatment in tumor therapy seems to have promising prospects. The current methods of treating tumors, including surgery, radiation therapy and chemotherapy, work well. As a novel biotherapy, mRNA therapy requires additional clinical trials to provide a better integration program before it can be effectively included in current procedures. In addition, mRNA technology may be utilized in conjunction with immunotherapy; currently, the majority of research is focused on immunomodulators. Owing to its distinct benefits, mRNA is also anticipated to supplant recombinant protein medications such as ILs, IFNs and cell colony factors. Tumor gene therapy may progress due to the significant ability of IVT mRNA to reduce miss effects in gene editing techniques. It is hypothesized that mRNA treatment may eventually lower the prevalence of hereditary family malignancies as tumor gene therapy is accomplished by introducing the target gene into the vector. Thus, a wider range of patients with cancer may benefit from these novel solutions.
Future research on mRNA vaccines for cancer should focus on the development of therapeutic and prophylactic vaccines. Personalized mRNA vaccines offer novel therapeutic strategies for tumors, such as pancreatic cancer, that do not respond to current treatments. In addition, mRNA technology can be used to develop vaccines for viruses that are important causes of cancer, such as HPV and HBV. A study indicated that HPV mRNA vaccines are more effective against tumors than HPV recombinant protein vaccines and HPV DNA vaccines in animal studies, supporting the future development of prophylactic vaccines against tumors using mRNA technology, which will be further evaluated in clinical trials (238).
Furthermore, the flexibility, adaptability and rapid production capacity of mRNA vaccines will offer exciting possibilities for the prevention and treatment of infectious diseases and cancers in the future, which will contribute to the reduction of the global disease burden.
Availability of data and materials
Not applicable.
Authors' contributions
BC, YY and XW wrote the manuscript; WY and EZ created figures and tables; YL, DW, YT and GT revised the manuscript; SS and JS supervised the research and revised the manuscript; KG supervised the research, revised the manuscript and provided financial support. All authors have read and agreed on the final version of the manuscript. Data authentication is not applicable.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Acknowledgements
Not applicable.
Funding
This study was supported by the Anhui Provincial Key Research and Development Project (grant no. 202004j07020044, chaired by KG).
References
|
Maomao C, He L, Dianqin S, Siyi H, Xinxin Y, Fan Y, Shaoli Z, Changfa X, Lin L, Ji P and Wanqing C: Current cancer burden in China: Epidemiology, etiology, and prevention. Cancer Biol Med. 19:1121–1138. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Roy PS and Saikia BJ: Cancer and cure: A critical analysis. Indian J Cancer. 53:441–442. 2016. View Article : Google Scholar | |
|
Sobhani N, Scaggiante B, Morris R, Chai D, Catalano M, Tardiel-Cyril DR, Neeli P, Roviello G, Mondani G and Li Y: Therapeutic cancer vaccines: From biological mechanisms and engineering to ongoing clinical trials. Cancer Treat Rev. 109:1024292022. View Article : Google Scholar : PubMed/NCBI | |
|
Corbett KS, F lynn B, Foulds K E, Francica J R, Boyoglu-Barnum S, Werner AP, Flach B, O'Connell S, Bock KW, Minai M, et al: Evaluation of the mRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. N Engl J Med. 383:1544–1555. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Scherubl H: Smoking tobacco and cancer risk. Dtsch Med Wochenschr. 146:412–417. 2021.In German. | |
|
van Elsland D and Neefjes J: Bacterial infections and cancer. EMBO Rep. 19:e466322018. View Article : Google Scholar : PubMed/NCBI | |
|
Schiller JT and Lowy DR: An introduction to virus infections and human cancer. Recent Results Cancer Res. 217:1–11. 2021. View Article : Google Scholar : | |
|
Zare Sakhvidi MJ, Lequy E, Goldberg M and Jacquemin B: Air pollution exposure and bladder, kidney and urinary tract cancer risk: A systematic review. Environ Pollut. 267:1153282020. View Article : Google Scholar : PubMed/NCBI | |
|
Yuan Y, Gao F, Chang Y, Zhao Q and He X: Advances of mRNA vaccine in tumor: A maze of opportunities and challenges. Biomark Res. 11:62023. View Article : Google Scholar : PubMed/NCBI | |
|
Gote V, Bolla PK, Kommineni N, Butreddy A, Nukala PK, Palakurthi SS and Khan W: A comprehensive review of mRNA Vaccines. Int J Mol Sci. 24:27002023. View Article : Google Scholar : PubMed/NCBI | |
|
Deng Z, Tian Y, Song J, An G and Yang P: mRNA Vaccines: The dawn of a new era of cancer immunotherapy. Front Immunol. 13:8871252022. View Article : Google Scholar : PubMed/NCBI | |
|
Liu C, Shi Q, Huang X, Koo S, Kong N and Tao W: mRNA-based cancer therapeutics. Nat Rev Cancer. 23:526–543. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Lorentzen CL, Haanen JB, Met Ö and Svane IM: Clinical advances and ongoing trials on mRNA vaccines for cancer treatment. Lancet Oncol. 23:e450–e458. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Weber JS, Carlino MS, Khattak A, Meniawy T, Ansstas G, Taylor MH, Kim KB, McKean M, Long GV, Sullivan RJ, et al: Individualised neoantigen therapy mRNA-4157 (V940) plus pembrolizumab versus pembrolizumab monotherapy in resected melanoma (KEYNOTE-942): A randomised, phase 2b study. Lancet. 403:632–644. 2024. View Article : Google Scholar : PubMed/NCBI | |
|
Siegel RL, Miller KD, Wagle NS and Jemal A: Cancer statistics, 2023. CA Cancer J Clin. 73:17–48. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Kaur R, Bhardwaj A and Gupta S: Cancer treatment therapies: Traditional to modern approaches to combat cancers. Mol Biol Rep. 50:9663–9676. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Sahin U and Tureci O: Personalized vaccines for cancer immunotherapy. Science. 359:1355–1360. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Saxena M, van der Burg SH, Melief CJM and Bhardwaj N: Therapeutic cancer vaccines. Nat Rev Cancer. 21:360–378. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Li Y, Wang M, Peng X, Yang Y, Chen Q, Liu J, She Q, Tan J, Lou C, Liao Z and Li X: mRNA vaccine in cancer therapy: Current advance and future outlook. Clin Transl Med. 13:e13842023. View Article : Google Scholar : PubMed/NCBI | |
|
Fang E, Liu X, Li M, Zhang Z, Song L, Zhu B, Wu X, Liu J, Zhao D and Li Y: Advances in COVID-19 mRNA vaccine development. Signal Transduct Target Ther. 7:942022. View Article : Google Scholar : PubMed/NCBI | |
|
Szabo GT, Mahiny AJ and Vlatkovic I: COVID-19 mRNA vaccines: Platforms and current developments. Mol Ther. 30:1850–1868. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Brenner S, Jacob F and Meselson M: An unstable intermediate carrying information from genes to ribosomes for protein synthesis. Nature. 190:576–581. 1961. View Article : Google Scholar : PubMed/NCBI | |
|
Malone RW, Felgner PL and Verma IM: Cationic liposome-mediated RNA transfection. Proc Natl Acad Sci USA. 86:6077–6081. 1989. View Article : Google Scholar : PubMed/NCBI | |
|
Wolff JA, Malone RW, Williams P, Chong W, Acsadi G, Jani A and Felgner PL: Direct gene transfer into mouse muscle in vivo. Science. 247(4949 Pt 1): 1465–1468. 1990. View Article : Google Scholar : PubMed/NCBI | |
|
Jirikowski GF, Sanna PP, Maciejewski-Lenoir D and Bloom FE: Reversal of diabetes insipidus in Brattleboro rats: Intrahypothalamic injection of vasopressin mRNA. Science. 255:996–998. 1992. View Article : Google Scholar : PubMed/NCBI | |
|
Martinon F, Krishnan S, Lenzen G, Magné R, Gomard E, Guillet JG, Lévy JP and Meulien P: Induction of virus-specific cytotoxic T lymphocytes in vivo by liposome-entrapped mRNA. Eur J Immunol. 23:1719–1722. 1993. View Article : Google Scholar : PubMed/NCBI | |
|
Boczkowski D, Nair SK, Snyder D and Gilboa E: Dendritic cells pulsed with RNA are potent antigen-presenting cells in vitro and in vivo. J Exp Med. 184:465–472. 1996. View Article : Google Scholar : PubMed/NCBI | |
|
Zhou WZ, Hoon DS, Huang SK, Fujii S, Hashimoto K, Morishita R and Kaneda Y: RNA melanoma vaccine: induction of antitumor immunity by human glycoprotein 100 mRNA immunization. Hum Gene Ther. 10:2719–2724. 1999. View Article : Google Scholar : PubMed/NCBI | |
|
Wang Y, Zhang Z, Luo J, Han X, Wei Y and Wei X: mRNA vaccine: A potential therapeutic strategy. Mol Cancer. 20:332021. View Article : Google Scholar : PubMed/NCBI | |
|
Wadhwa A, Aljabbari A, Lokras A, Foged C and Thakur A: Opportunities and Challenges in the Delivery of mRNA-based Vaccines. Pharmaceutics. 12:1022020. View Article : Google Scholar : PubMed/NCBI | |
|
Kackos CM, Surman SL, Jones BG, Sealy RE, Jeevan T, Davitt CJH, Pustylnikov S, Darling TL, Boon ACM, Hurwitz JL, et al: mRNA Vaccine Mitigates SARS-CoV-2 Infections and COVID-19. Microbiol Spectr. 11:e04240222023. View Article : Google Scholar : PubMed/NCBI | |
|
Park JW, Lagniton PNP, Liu Y and Xu RH: mRNA vaccines for COVID-19: What, why and how. Int J Biol Sci. 17:1446–1460. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Gong H, Wen J, Luo R, Feng Y, Guo J, Fu H and Zhou X: Integrated mRNA sequence optimization using deep learning. Brief Bioinform. 24:bbad0012023. View Article : Google Scholar : PubMed/NCBI | |
|
Pardi N, Hogan MJ, Porter FW and Weissman D: mRNA vaccines-a new era in vaccinology. Nat Rev Drug Discov. 17:261–279. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Herrera M, Kim J, Eygeris Y, Jozic A and Sahay G: Illuminating endosomal escape of polymorphic lipid nanoparticles that boost mRNA delivery. Biomater Sci. 9:4289–4300. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Jyotsana N, Sharma A, Chaturvedi A, Budida R, Scherr M, Kuchenbauer F, Lindner R, Noyan F, Sühs KW, Stangel M, et al: Lipid nanoparticle-mediated siRNA delivery for safe targeting of human CML in vivo. Ann Hematol. 98:1905–1918. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Sahin U, Kariko K and Tureci O: mRNA-based therapeutics-developing a new class of drugs. Nat Rev Drug Discov. 13:759–780. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Cruz FM, Chan A and Rock KL: Pathways of MHC I cross-presentation of exogenous antigens. Semin Immunol. 66:1017292023. View Article : Google Scholar : PubMed/NCBI | |
|
Rosa SS, Prazeres DMF, Azevedo AM and Marques MPC: mRNA vaccines manufacturing: Challenges and bottlenecks. Vaccine. 39:2190–2200. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Xu S, Yang K, Li R and Zhang L: mRNA Vaccine Era-Mechanisms, drug platform and clinical prospection. Int J Mol Sci. 21:20202020. View Article : Google Scholar | |
|
Talotta R: Do COVID-19 RNA-based vaccines put at risk of immune-mediated diseases? In reply to 'potential antigenic cross-reactivity between SARS-CoV-2 and human tissue with a possible link to an increase in autoimmune diseases'. Clin Immunol. 224:1086652021. View Article : Google Scholar | |
|
Chen YG and Hur S: Cellular origins of dsRNA, their recognition and consequences. Nat Rev Mol Cell Biol. 23:286–301. 2022. View Article : Google Scholar : | |
|
Vaidyanathan S, Azizian KT, Haque AKMA, Henderson JM, Hendel A, Shore S, Antony JS, Hogrefe RI, Kormann MSD, Porteus MH and McCaffrey AP: Uridine depletion and chemical modification increase Cas9 mRNA activity and reduce immunogenicity without HPLC purification. Mol Ther Nucleic Acids. 12:530–542. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Bernardo M, Tolstykh T, Zhang YA, Bangari DS, Cao H, Heyl KA, Lee JS, Malkova NV, Malley K, Marquez E, et al: An experimental model of anti-PD-1 resistance exhibits activation of TGFβ and Notch pathways and is sensitive to local mRNA immunotherapy. Oncoimmunology. 10:18812682021. View Article : Google Scholar | |
|
Yao R, Xie C and Xia X: Recent progress in mRNA cancer vaccines. Hum Vaccin Immunother. 20:23071872024. View Article : Google Scholar : PubMed/NCBI | |
|
Wilson RC and Carroll D: The daunting economics of therapeutic genome editing. CRISPR J. 2:280–284. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Morrison C: $1-million price tag set for Glybera gene therapy. Nat Biotechnol. 33:217–218. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Jackson NAC, Kester KE, Casimiro D, Gurunathan S and DeRosa F: The promise of mRNA vaccines: A biotech and industrial perspective. NPJ Vaccines. 5:112020. View Article : Google Scholar : PubMed/NCBI | |
|
Miao L, Zhang Y and Huang L: mRNA vaccine for cancer immunotherapy. Mol Cancer. 20:412021. View Article : Google Scholar : PubMed/NCBI | |
|
Ita K: Coronavirus Disease (COVID-19): Current status and prospects for drug and vaccine development. Arch Med Res. 52:15–24. 2021. View Article : Google Scholar : | |
|
Van Nuffel AM, Wilgenhof S, Thielemans K and Bonehill A: Overcoming HLA restriction in clinical trials: Immune monitoring of mRNA-loaded DC therapy. Oncoimmunology. 1:1392–1394. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Ho W, Gao M, Li F, Li Z, Zhang XQ and Xu X: Next-Generation Vaccines: Nanoparticle-Mediated DNA and mRNA Delivery. Adv Healthc Mater. 10:e20018122021. View Article : Google Scholar : PubMed/NCBI | |
|
Guevara ML, Persano F and Persano S: Advances in lipid nanoparticles for mRNA-Based cancer immunotherapy. Front Chem. 8:5899592020. View Article : Google Scholar : PubMed/NCBI | |
|
De Beuckelaer A, Grooten J and De Koker S: Type I Interferons Modulate CD8(+) T Cell Immunity to mRNA Vaccines. Trends Mol Med. 23:216–226. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Karam M and Daoud G: mRNA vaccines: Past, present, future. Asian J Pharm Sci. 17:491–522. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Zhao P, Hou X, Yan J, Du S, Xue Y, Li W, Xiang G and Dong Y: Long-term storage of lipid-like nanoparticles for mRNA delivery. Bioact Mater. 5:358–363. 2020.PubMed/NCBI | |
|
Stitz L, Vogel A, Schnee M, Voss D, Rauch S, Mutzke T, Ketterer T, Kramps T and Petsch B: A thermostable messenger RNA based vaccine against rabies. PLoS Negl Trop Dis. 11:e00061082017. View Article : Google Scholar : PubMed/NCBI | |
|
Petsch B, Schnee M, Vogel AB, Lange E, Hoffmann B, Voss D, Schlake T, Thess A, Kallen KJ, Stitz L and Kramps T: Protective efficacy of in vitro synthesized, specific mRNA vaccines against influenza A virus infection. Nat Biotechnol. 30:1210–1216. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Dowdy SF: Overcoming cellular barriers for RNA therapeutics. Nat Biotechnol. 35:222–229. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Chaudhary N, Weissman D and Whitehead KA: mRNA vaccines for infectious diseases: principles, delivery and clinical translation. Nat Rev Drug Discov. 20:817–838. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Heine A, Juranek S and Brossart P: Clinical and immunological effects of mRNA vaccines in malignant diseases. Mol Cancer. 20:522021. View Article : Google Scholar : PubMed/NCBI | |
|
Schlake T, Thess A, Fotin-Mleczek M and Kallen KJ: Developing mRNA-vaccine technologies. RNA Biol. 9:1319–1330. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Dobrowolski C, Paunovska K, Schrader Echeverri E, Loughrey D, Da Silva Sanchez AJ, Ni H, Hatit MZC, Lokugamage MP, Kuzminich Y, Peck HE, et al: Nanoparticle single-cell multiomic readouts reveal that cell heterogeneity influences lipid nanoparticle-mediated messenger RNA delivery. Nat Nanotechnol. 17:871–879. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Solodushko V and Fouty B: Terminal hairpins improve protein expression in IRES-initiated mRNA in the absence of a cap and polyadenylated tail. Gene Ther. 30:620–627. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Kowalski PS, Rudra A, Miao L and Anderson DG: Delivering the messenger: Advances in technologies for therapeutic mRNA delivery. Mol Ther. 27:710–728. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Park KS, Sun X, Aikins ME and Moon JJ: Non-viral COVID-19 vaccine delivery systems. Adv Drug Deliv Rev. 169:137–151. 2021. View Article : Google Scholar | |
|
Kim J, Eygeris Y, Gupta M and Sahay G: Self-assembled mRNA vaccines. Adv Drug Deliv Rev. 170:83–112. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Kauffman KJ, Webber MJ and Anderson DG: Materials for non-viral intracellular delivery of messenger RNA therapeutics. J Control Release. 240:227–234. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Whitley J, Zwolinski C, Denis C, Maughan M, Hayles L, Clarke D, Snare M, Liao H, Chiou S, Marmura T, et al: Development of mRNA manufacturing for vaccines and therapeutics: mRNA platform requirements and development of a scalable production process to support early phase clinical trials. Transl Res. 242:38–55. 2022. View Article : Google Scholar | |
|
Kariko K, Muramatsu H, Ludwig J and Weissman D: Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. 39:e1422011. View Article : Google Scholar : PubMed/NCBI | |
|
Linares-Fernandez S, Lacroix C, Exposito JY and Verrier B: Tailoring mRNA vaccine to balance innate/adaptive immune response. Trends Mol Med. 26:311–323. 2020. View Article : Google Scholar | |
|
Engel BJ, Grindel BJ, Gray JP and Millward SW: Purification of poly-dA oligonucleotides and mRNA-protein fusions with dT(25)-OAS resin. Bioorg Med Chem Lett. 30:1269342020. View Article : Google Scholar : PubMed/NCBI | |
|
Ye Z, Harmon J, Ni W, Li Y, Wich D and Xu Q: The mRNA Vaccine Revolution: COVID-19 has launched the future of vaccinology. ACS Nano. 17:15231–15253. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
To KKW and Cho WCS: An overview of rational design of mRNA-based therapeutics and vaccines. Expert Opin Drug Discov. 16:1307–1317. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Kariko K, Kuo A and Barnathan E: Overexpression of urokinase receptor in mammalian cells following administration of the in vitro transcribed encoding mRNA. Gene Ther. 6:1092–1100. 1999. View Article : Google Scholar : PubMed/NCBI | |
|
Kallen KJ and Theβ A: A development that may evolve into a revolution in medicine: mRNA as the basis for novel, nucleotide-based vaccines and drugs. Ther Adv Vaccines. 2:10–31. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Holtkamp S, Kreiter S, Selmi A, Simon P, Koslowski M, Huber C, Türeci O and Sahin U: Modification of antigen-encoding RNA increases stability, translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood. 108:4009–4017. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Leppek K, Byeon GW, Kladwang W, Wayment-Steele HK, Kerr CH, Xu AF, Kim DS, Topkar VV, Choe C, Rothschild D, et al: Combinatorial optimization of mRNA structure, stability, and translation for RNA-based therapeutics. Nat Commun. 13:15362022. View Article : Google Scholar : PubMed/NCBI | |
|
Ramanathan A, Robb GB and Chan SH: mRNA capping: Biological functions and applications. Nucleic Acids Res. 44:7511–7526. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Li Y and Kiledjian M: Regulation of mRNA decapping. Wiley Interdiscip Rev RNA. 1:253–265. 2010. View Article : Google Scholar | |
|
Geall AJ, Verma A, Otten GR, Shaw CA, Hekele A, Banerjee K, Cu Y, Beard CW, Brito LA, Krucker T, et al: Nonviral delivery of self-amplifying RNA vaccines. Proc Natl Acad Sci USA. 109:14604–14609. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Mohamad Razif MI, Nizar N, Zainal Abidin NH, Muhammad Ali SN, Wan Zarimi WNN, Khotib J, Susanti D, Mohd Jailani MT and Taher M: Emergence of mRNA vaccines in the management of cancer. Expert Rev Vaccines. 22:629–642. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Kim SC, Sekhon SS, Shin WR, Ahn G, Cho BK, Ahn JY and Kim YH: Modifications of mRNA vaccine structural elements for improving mRNA stability and translation efficiency. Mol Cell Toxicol. 18:1–8. 2022. View Article : Google Scholar | |
|
Warminski M, Kowalska J, Nowak E, Kubacka D, Tibble R, Kasprzyk R, Sikorski PJ, Gross JD, Nowotny M and Jemielity J: Structural Insights into the interaction of clinically relevant phosphorothioate mRNA Cap Analogs with translation initiation factor 4E Reveal Stabilization via Electrostatic Thio-Effect. ACS Chem Biol. 16:334–343. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Kuhn AN, Diken M, Kreiter S, Selmi A, Kowalska J, Jemielity J, Darzynkiewicz E, Huber C, Türeci O and Sahin U: Phosphorothioate cap analogs increase stability and translational efficiency of RNA vaccines in immature dendritic cells and induce superior immune responses in vivo. Gene Ther. 17:961–971. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Grudzien-Nogalska E, Jemielity J, Kowalska J, Darzynkiewicz E and Rhoads RE: Phosphorothioate cap analogs stabilize mRNA and increase translational efficiency in mammalian cells. RNA. 13:1745–1755. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Rydzik AM, Kulis M, Lukaszewicz M, Kowalska J, Zuberek J, Darzynkiewicz ZM, Darzynkiewicz E and Jemielity J: Synthesis and properties of mRNA cap analogs containing imidodiphosphate moiety-fairly mimicking natural cap structure, yet resistant to enzymatic hydrolysis. Bioorg Med Chem. 20:1699–1710. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Henderson JM, Ujita A, Hill E, Yousif-Rosales S, Smith C, Ko N, McReynolds T, Cabral CR, Escamilla-Powers JR and Houston ME: Cap 1 Messenger RNA Synthesis with Co-transcriptional CleanCap((R)) Analog by In Vitro Transcription. Curr Protoc. 1:e392021. View Article : Google Scholar | |
|
Suknuntha K, Tao L, Brok-Volchanskaya V, D'Souza SS, Kumar A and Slukvin I: Optimization of Synthetic mRNA for highly efficient translation and its application in the generation of endothelial and hematopoietic cells from human and primate pluripotent stem cells. Stem Cell Rev Rep. 14:525–534. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Ryczek N, Lys A and Makalowska I: The Functional Meaning of 5'UTR in Protein-Coding Genes. Int J Mol Sci. 24:29762023. View Article : Google Scholar : PubMed/NCBI | |
|
Creusot RJ, Chang P, Healey DG, Tcherepanova IY, Nicolette CA and Fathman CG: A short pulse of IL-4 delivered by DCs electroporated with modified mRNA can both prevent and treat autoimmune diabetes in NOD mice. Mol Ther. 18:2112–2120. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Asrani KH, Farelli JD, Stahley MR, Miller RL, Cheng CJ, Subramanian RR and Brown JM: Optimization of mRNA untranslated regions for improved expression of therapeutic mRNA. RNA Biol. 15:756–762. 2018.PubMed/NCBI | |
|
Chakraborty C, Sharma AR, Bhattacharya M and Lee SS: From COVID-19 to Cancer mRNA Vaccines: Moving from bench to clinic in the vaccine landscape. Front Immunol. 12:6793442021. View Article : Google Scholar : PubMed/NCBI | |
|
Beaudoin CA, Bartas M, Volná A, Pečinka P and Blundell TL: Are There Hidden Genes in DNA/RNA Vaccines? Front Immunol. 13:8019152022. View Article : Google Scholar : PubMed/NCBI | |
|
Gustafsson C, Govindarajan S and Minshull J: Codon bias and heterologous protein expression. Trends Biotechnol. 22:346–353. 2004. View Article : Google Scholar : PubMed/NCBI | |
|
Passmore LA and Coller J: Roles of mRNA poly(A) tails in regulation of eukaryotic gene expression. Nat Rev Mol Cell Biol. 23:93–106. 2022. View Article : Google Scholar | |
|
Lima SA, Chipman LB, Nicholson AL, Chen YH, Yee BA, Yeo GW, Coller J and Pasquinelli AE: Short poly(A) tails are a conserved feature of highly expressed genes. Nat Struct Mol Biol. 24:1057–1063. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
McNamara MA, Nair SK and Holl EK: RNA-Based Vaccines in Cancer Immunotherapy. J Immunol Res. 2015:7945282015. View Article : Google Scholar : PubMed/NCBI | |
|
Chung JY, Thone MN and Kwon YJ: COVID-19 vaccines: The status and perspectives in delivery points of view. Adv Drug Deliv Rev. 170:1–25. 2021. View Article : Google Scholar | |
|
Islam MA, Rice J, Reesor E, Zope H, Tao W, Lim M, Ding J, Chen Y, Aduluso D, Zetter BR, et al: Adjuvant-pulsed mRNA vaccine nanoparticle for immunoprophylactic and therapeutic tumor suppression in mice. Biomaterials. 266:1204312021. View Article : Google Scholar | |
|
De Keersmaecker B, Claerhout S, Carrasco J, Bar I, Corthals J, Wilgenhof S, Neyns B and Thielemans K: TriMix and tumor antigen mRNA electroporated dendritic cell vaccination plus ipilimumab: link between T-cell activation and clinical responses in advanced melanoma. J Immunother Cancer. 8:e0003292020. View Article : Google Scholar : PubMed/NCBI | |
|
Franzoni G, Anfossi A, De Ciucis CG, Mecocci S, Carta T, Dei Giudici S, Fruscione F, Zinellu S, Vito G, Graham SP, et al: Targeting Toll-Like Receptor 2: Polarization of Porcine Macrophages by a Mycoplasma-Derived Pam2cys Lipopeptide. Vaccines (Basel). 9:6922021. View Article : Google Scholar : PubMed/NCBI | |
|
Gu Y, Yang J, He C, Zhao T, Lu R, Liu J, Mo X, Wen F and Shi H: Incorporation of a Toll-like receptor 2/6 agonist potentiates mRNA vaccines against cancer and infectious diseases. Signal Transduct Target Ther. 8:2732023. View Article : Google Scholar : PubMed/NCBI | |
|
Ulmer JB, Mason PW, Geall A and Mandl CW: RNA-based vaccines. Vaccine. 30:4414–4418. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
McCullough KC, Milona P, Thomann-Harwood L, Démoulins T, Englezou P, Suter R and Ruggli N: Self-Amplifying Replicon RNA Vaccine Delivery to Dendritic Cells by Synthetic Nanoparticles. Vaccines (Basel). 2:735–754. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Maruggi G, Zhang C, Li J, Ulmer JB and Yu D: mRNA as a transformative technology for vaccine development to control infectious diseases. Mol Ther. 27:757–772. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Zhuang X, Qi Y, Wang M, Yu N, Nan F, Zhang H, Tian M, Li C, Lu H and Jin N: mRNA Vaccines Encoding the HA Protein of Influenza A H1N1 virus delivered by cationic lipid nanoparticles induce protective immune responses in mice. Vaccines (Basel). 8:1232020. View Article : Google Scholar : PubMed/NCBI | |
|
Kallen KJ, Heidenreich R, Schnee M, Petsch B, Schlake T, Thess A, Baumhof P, Scheel B, Koch SD and Fotin-Mleczek M: A novel, disruptive vaccination technology: Self-adjuvanted RNActive((R)) vaccines. Hum Vaccin Immunother. 9:2263–2276. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Leal L, Guardo AC, Morón-López S, Salgado M, Mothe B, Heirman C, Pannus P, Vanham G, van den Ham HJ, Gruters R, et al: Phase I clinical trial of an intranodally administered mRNA-based therapeutic vaccine against HIV-1 infection. AIDS. 32:2533–2545. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Jong W, Leal L, Buyze J, Pannus P, Guardo A, Salgado M, Mothe B, Molto J, Moron-Lopez S, Gálvez C, et al: Therapeutic Vaccine in Chronically HIV-1-Infected Patients: A Randomized, Double-Blind, Placebo-Controlled Phase IIa Trial with HTI-TriMix. Vaccines (Basel). 7:2092019. View Article : Google Scholar : PubMed/NCBI | |
|
Egan KP, Hook LM, Naughton A, Pardi N, Awasthi S, Cohen GH, Weissman D and Friedman HM: An HSV-2 nucleoside-modified mRNA genital herpes vaccine containing glycoproteins gC, gD, and gE protects mice against HSV-1 genital lesions and latent infection. PLoS Pathog. 16:e10087952020. View Article : Google Scholar : PubMed/NCBI | |
|
Wollner CJ, Richner M, Hassert MA, Pinto AK, Brien JD and Richner JM: A Dengue Virus Serotype 1 mRNA-LNP vaccine elicits protective immune responses. J Virol. 95:e02482–20. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Wollner CJ and Richner JM: mRNA Vaccines against Flaviviruses. Vaccines (Basel). 9:1482021. View Article : Google Scholar | |
|
Rzymski P, Szuster-Ciesielska A, Dzieciątkowski T, Gwenzi W and Fal A: mRNA vaccines: The future of prevention of viral infections? J Med Virol. 95:e285722023. View Article : Google Scholar : PubMed/NCBI | |
|
Richner JM, Himansu S, Dowd KA, Butler SL, Salazar V, Fox JM, Julander JG, Tang WW, Shresta S, Pierson TC, et al: Modified mRNA Vaccines Protect against Zika Virus Infection. Cell. 169:1762017. View Article : Google Scholar : PubMed/NCBI | |
|
Parums DV: Editorial: mRNA Vaccines and Future Epidemic, Pandemic, and Endemic Zoonotic Virus Infections. Med Sci Monit. 27:e9329152021.PubMed/NCBI | |
|
Haynes BF: A new vaccine to battle covid-19. N Engl J Med. 384:470–471. 2021. View Article : Google Scholar | |
|
Fairweather D, Beetler DJ, Di Florio DN, Musigk N, Heidecker B and Cooper LT Jr: COVID-19, myocarditis and pericarditis. Circ Res. 132:1302–1319. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Maruggi G, Chiarot E, Giovani C, Buccato S, Bonacci S, Frigimelica E, Margarit I, Geall A, Bensi G and Maione D: Immunogenicity and protective efficacy induced by self-amplifying mRNA vaccines encoding bacterial antigens. Vaccine. 35:361–368. 2017. View Article : Google Scholar | |
|
Jawalagatti V, Kirthika P and Lee JH: Oral mRNA Vaccines Against Infectious Diseases-A Bacterial Perspective (Invited). Front Immunol. 13:8848622022. View Article : Google Scholar | |
|
Beck JD, Reidenbach D, Salomon N, Sahin U, Türeci Ö, Vormehr M and Kranz LM: mRNA therapeutics in cancer immunotherapy. Mol Cancer. 20:692021. View Article : Google Scholar : PubMed/NCBI | |
|
Vishweshwaraiah YL and Dokholyan NV: mRNA vaccines for cancer immunotherapy. Front Immunol. 13:10290692022. View Article : Google Scholar : | |
|
Wang F: Identification of tumor antigens and immune subtypes of acute myeloid leukemia for mRNA vaccine development. Clin Transl Oncol. 25:2204–2223. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Chung DJ, Sharma S, Rangesa M, DeWolf S, Elhanati Y, Perica K and Young JW: Langerhans dendritic cell vaccine bearing mRNA-encoded tumor antigens induces antimyeloma immunity after autotransplant. Blood Adv. 6:1547–1558. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Wang S, Yang Y, Li L, Ma P, Jiang Y, Ge M, Yu Y, Huang H, Fang Y, Jiang N, et al: Identification of Tumor Antigens and Immune Subtypes of Malignant Mesothelioma for mRNA Vaccine Development. Vaccines (Basel). 10:11682022. View Article : Google Scholar : PubMed/NCBI | |
|
Lin H, Wang K, Xiong Y, Zhou L, Yang Y, Chen S, Xu P, Zhou Y, Mao R, Lv G, et al: Identification of tumor antigens and immune subtypes of glioblastoma for mRNA vaccine development. Front Immunol. 13:7732642022. View Article : Google Scholar : PubMed/NCBI | |
|
Zhou Q, Yan X, Zhu H, Xin Z, Zhao J, Shen W, Yin W, Guo Y, Xu H, Zhao M, et al: Identification of three tumor antigens and immune subtypes for mRNA vaccine development in diffuse glioma. Theranostics. 11:9775–9790. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Xu H, Zheng X, Zhang S, Yi X, Zhang T, Wei Q, Li H and Ai J: Tumor antigens and immune subtypes guided mRNA vaccine development for kidney renal clear cell carcinoma. Mol Cancer. 20:1592021. View Article : Google Scholar : PubMed/NCBI | |
|
Rojas LA, Sethna Z, Soares KC, Olcese C, Pang N, Patterson E, Lihm J, Ceglia N, Guasp P, Chu A, et al: Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature. 618:144–150. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
mRNA Vaccine Slows Melanoma Recurrence. Cancer Discov. 13:12782023. View Article : Google Scholar | |
|
Sun Z, Jing C, Zhan H, Guo X, Suo N, Kong F, Tao W, Xiao C, Hu D, Wang H and Jiang S: Identification of tumor antigens and immune landscapes for bladder urothelial carcinoma mRNA vaccine. Front Immunol. 14:10974722023. View Article : Google Scholar : PubMed/NCBI | |
|
Kiousi E, Lyraraki V, Mardiki GL, Stachika N, Damianou AK, Malainou CP, Syrigos N, Gomatou G and Kotteas E: Progress and Challenges of Messenger RNA Vaccines in the Therapeutics of NSCLC. Cancers (Basel). 15:55892023. View Article : Google Scholar : PubMed/NCBI | |
|
Zheng X, Xu H, Yi X, Zhang T, Wei Q, Li H and Ai J: Tumor-antigens and immune landscapes identification for prostate adenocarcinoma mRNA vaccine. Mol Cancer. 20:1602021. View Article : Google Scholar : PubMed/NCBI | |
|
Wu Y, Li Z, Lin H and Wang H: Identification of tumor antigens and immune subtypes of high-grade serous ovarian cancer for mRNA vaccine development. J Cancer. 14:2655–2669. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Song J, Zhang Y, Zhou C, Zhan J, Cheng X, Huang H, Mao S and Zong Z: The dawn of a new Era: mRNA vaccines in colorectal cancer immunotherapy. Int Immunopharmacol. 132:1120372024. View Article : Google Scholar : PubMed/NCBI | |
|
Sumi T, Koshino Y, Michimata H, Nagayama D, Watanabe H, Yamada Y and Chiba H: Cytokine release syndrome in a patient with non-small cell lung cancer on ipilimumab and nivolumab maintenance therapy after vaccination with the mRNA-1273 vaccine: A case report. Transl Lung Cancer Res. 11:1973–1976. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Rausch S, Schwentner C, Stenzl A and Bedke J: mRNA vaccine CV9103 and CV9104 for the treatment of prostate cancer. Hum Vaccin Immunother. 10:3146–3152. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Liu L, Wang Y, Miao L, Liu Q, Musetti S, Li J and Huang L: Combination Immunotherapy of MUC1 mRNA Nano-vaccine and CTLA-4 Blockade Effectively Inhibits Growth of Triple Negative Breast Cancer. Mol Ther. 26:45–55. 2018. View Article : Google Scholar : | |
|
Liu C, Papukashvili D, Dong Y, Wang X, Hu X, Yang N, Cai J, Xie F, Rcheulishvili N and Wang PG: Identification of Tumor Antigens and Design of mRNA vaccine for colorectal cancer based on the immune subtype. Front Cell Dev Biol. 9:7835272022. View Article : Google Scholar : PubMed/NCBI | |
|
Huang X, Zhang G, Tang TY, Gao X and Liang TB: Personalized pancreatic cancer therapy: From the perspective of mRNA vaccine. Mil Med Res. 9:532022.PubMed/NCBI | |
|
Samuel N and Hudson TJ: The molecular and cellular heterogeneity of pancreatic ductal adenocarcinoma. Nat Rev Gastroenterol Hepatol. 9:77–87. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Makohon-Moore AP, Zhang M, Reiter JG, Bozic I, Allen B, Kundu D, Chatterjee K, Wong F, Jiao Y, Kohutek ZA, et al: Limited heterogeneity of known driver gene mutations among the metastases of individual patients with pancreatic cancer. Nat Genet. 49:358–366. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Bear AS, Vonderheide RH and O'Hara MH: Challenges and opportunities for pancreatic cancer immunotherapy. Cancer Cell. 38:788–802. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Kang N, Zhang S and Wang Y: A personalized mRNA vaccine has exhibited potential in the treatment of pancreatic cancer. Holist Integr Oncol. 2:182023. View Article : Google Scholar : PubMed/NCBI | |
|
Tan H, Yu T, Liu C, Wang Y, Jing F, Ding Z, Liu J and Shi H: Identifying tumor antigens and immuno-subtyping in colon adenocarcinoma to facilitate the development of mRNA vaccine. Cancer Med. 11:4656–4672. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Ping H, Yu W, Gong X, Tong X, Lin C, Chen Z, Cai C, Guo K and Ke H: Analysis of melanoma tumor antigens and immune subtypes for the development of mRNA vaccine. Invest New Drugs. 40:1173–1184. 2022. View Article : Google Scholar : | |
|
Sittplangkoon C, Alameh MG, Weissman D, Lin PJC, Tam YK, Prompetchara E and Palaga T: mRNA vaccine with unmodified uridine induces robust type I interferon-dependent anti-tumor immunity in a melanoma model. Front Immunol. 13:9830002022. View Article : Google Scholar : | |
|
Li RQ, Wang W, Yan L, Song LY, Guan X, Zhang W and Lian J: Identification of tumor antigens and immune subtypes in breast cancer for mRNA vaccine development. Front Oncol. 12:9737122022. View Article : Google Scholar : PubMed/NCBI | |
|
Niemi JVL, Sokolov AV and Schioth HB: Neoantigen vaccines; clinical trials, classes, indications, adjuvants and combinatorial treatments. Cancers (Basel). 14:51632022. View Article : Google Scholar : PubMed/NCBI | |
|
Oberli MA, Reichmuth AM, Dorkin JR, Mitchell MJ, Fenton OS, Jaklenec A, Anderson DG, Langer R and Blankschtein D: Lipid Nanoparticle Assisted mRNA delivery for potent cancer immunotherapy. Nano Lett. 17:1326–1335. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Pardoll DM: The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer. 12:252–264. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Mellor AL, Keskin DB, Johnson T, Chandler P and Munn DH: Cells expressing indoleamine 2,3-dioxygenase inhibit T cell responses. J Immunol. 168:3771–3776. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Mellman I, Coukos G and Dranoff G: Cancer immunotherapy comes of age. Nature. 480:480–489. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Conry RM, LoBuglio AF, Wright M, Sumerel L, Pike MJ, Johanning F, Benjamin R, Lu D and Curiel DT: Characterization of a messenger RNA polynucleotide vaccine vector. Cancer Res. 55:1397–1400. 1995.PubMed/NCBI | |
|
Huang X, Tang T, Zhang G and Liang T: Identification of tumor antigens and immune subtypes of cholangiocarcinoma for mRNA vaccine development. Mol Cancer. 20:502021. View Article : Google Scholar : | |
|
Guo Y, Lei K and Tang L: Neoantigen Vaccine delivery for personalized anticancer immunotherapy. Front Immunol. 9:14992018. View Article : Google Scholar : PubMed/NCBI | |
|
Grabbe S, Haas H, Diken M, Kranz LM, Langguth P and Sahin U: Translating nanoparticulate-personalized cancer vaccines into clinical applications: Case study with RNA-lipoplexes for the treatment of melanoma. Nanomedicine (Lond). 11:2723–2734. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Esprit A, de Mey W, Bahadur Shahi R, Thielemans K, Franceschini L and Breckpot K: Neo-Antigen mRNA Vaccines. Vaccines (Basel). 8:7762020. View Article : Google Scholar : PubMed/NCBI | |
|
Blass E and Ott PA: Advances in the development of personalized neoantigen-based therapeutic cancer vaccines. Nat Rev Clin Oncol. 18:215–229. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Cafri G, Gartner JJ, Zaks T, Hopson K, Levin N, Paria BC, Parkhurst MR, Yossef R, Lowery FJ and Jafferji MS: mRNA vaccine-induced neoantigen-specific T cell immunity in patients with gastrointestinal cancer. J Clin Invest. 130:5976–5988. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Roberts ND, Kortschak RD, Parker WT, Schreiber AW, Branford S, Scott HS, Glonek G and Adelson DL: A comparative analysis of algorithms for somatic SNV detection in cancer. Bioinformatics. 29:2223–2230. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Karasaki T, Nagayama K, Kuwano H, Nitadori JI, Sato M, Anraku M, Hosoi A, Matsushita H, Takazawa M, Ohara O, et al: Prediction and prioritization of neoantigens: integration of RNA sequencing data with whole-exome sequencing. Cancer Sci. 108:170–177. 2017. View Article : Google Scholar : | |
|
Horak P, Frohling S and Glimm H: Integrating next-generation sequencing into clinical oncology: strategies, promises and pitfalls. ESMO Open. 1:e0000942016. View Article : Google Scholar : PubMed/NCBI | |
|
Bassani-Sternberg M, Bräunlein E, Klar R, Engleitner T, Sinitcyn P, Audehm S, Straub M, Weber J, Slotta-Huspenina J, Specht K, et al: Direct identification of clinically relevant neoepitopes presented on native human melanoma tissue by mass spectrometry. Nat Commun. 7:134042016. View Article : Google Scholar : PubMed/NCBI | |
|
Kongsted P, Borch TH, Ellebaek E, Iversen TZ, Andersen R, Met Ö, Hansen M, Lindberg H, Sengeløv L and Svane IM: Dendritic cell vaccination in combination with docetaxel for patients with metastatic castration-resistant prostate cancer: A randomized phase II study. Cytotherapy. 19:500–513. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Ingels J, De Cock L, Mayer RL, Devreker P, Weening K, Heyns K, Lootens N, De Smet S, Brusseel M, De Munter S, et al: Small-scale manufacturing of neoantigen-encoding messenger RNA for early-phase clinical trials. Cytotherapy. 24:213–222. 2022. View Article : Google Scholar | |
|
Sahin U, Derhovanessian E, Miller M, Kloke BP, Simon P, Löwer M, Bukur V, Tadmor AD, Luxemburger U, Schrörs B, et al: Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature. 547:222–226. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Steitz J, Britten CM, Wölfel T and Tüting T: Effective induction of anti-melanoma immunity following genetic vaccination with synthetic mRNA coding for the fusion protein EGFP.TRP2. Cancer Immunol Immunother. 55:246–253. 2006. View Article : Google Scholar | |
|
Weide B, Pascolo S, Scheel B, Derhovanessian E, Pflugfelder A, Eigentler TK, Pawelec G, Hoerr I, Rammensee HG and Garbe C: Direct injection of protamine-protected mRNA: results of a phase 1/2 vaccination trial in metastatic melanoma patients. J Immunother. 32:498–507. 2009. View Article : Google Scholar | |
|
Mai Y, Guo J, Zhao Y, Ma S, Hou Y and Yang J: Intranasal delivery of cationic liposome-protamine complex mRNA vaccine elicits effective anti-tumor immunity. Cell Immunol. 354:1041432020. View Article : Google Scholar | |
|
Sahin U, Oehm P, Derhovanessian E, Jabulowsky RA, Vormehr M, Gold M, Maurus D, Schwarck-Kokarakis D, Kuhn AN, Omokoko T, et al: An RNA vaccine drives immunity in checkpoint-inhibitor-treated melanoma. Nature. 585:107–112. 2020. View Article : Google Scholar | |
|
Conlon KC, Miljkovic MD and Waldmann TA: Cytokines in the Treatment of Cancer. J Interferon Cytokine Res. 39:6–21. 2019. View Article : Google Scholar : | |
|
Di Trani CA, Fernandez-Sendin M, Cirella A, Segués A, Olivera I, Bolaños E, Melero I and Berraondo P: Advances in mRNA-based drug discovery in cancer immunotherapy. Expert Opin Drug Discov. 17:41–53. 2022. View Article : Google Scholar | |
|
Komel T, Bosnjak M, Kranjc Brezar S, De Robertis M, Mastrodonato M, Scillitani G, Pesole G, Signori E, Sersa G and Cemazar M: Gene electrotransfer of IL-2 and IL-12 plasmids effectively eradicated murine B16.F10 melanoma. Bioelectrochemistry. 141:1078432021. View Article : Google Scholar : PubMed/NCBI | |
|
Etxeberria I, Bolaños E, Quetglas JI, Gros A, Villanueva A, Palomero J, Sánchez-Paulete AR, Piulats JM, Matias-Guiu X, Olivera I, et al: Intratumor Adoptive Transfer of IL-12 mRNA Transiently Engineered Antitumor CD8(+) T Cells. Cancer Cell. 36:613–629 e7. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Hewitt SL, Bailey D, Zielinski J, Apte A, Musenge F, Karp R, Burke S, Garcon F, Mishra A, Gurumurthy S, et al: Intratumoral IL12 mRNA Therapy Promotes TH1 transformation of the tumor microenvironment. Clin Cancer Res. 26:6284–6298. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Hotz C, Wagenaar TR, Gieseke F, Bangari DS, Callahan M, Cao H, Diekmann J, Diken M, Grunwitz C, Hebert A, et al: Local delivery of mRNA-encoded cytokines promotes antitumor immunity and tumor eradication across multiple preclinical tumor models. Sci Transl Med. 13:eabc78042021. View Article : Google Scholar : PubMed/NCBI | |
|
Hewitt SL, Bai A, Bailey D, Ichikawa K, Zielinski J, Karp R, Apte A, Arnold K, Zacharek SJ, Iliou MS, et al: Durable anticancer immunity from intratumoral administration of IL-23, IL-36ү, and OX40L mRNAs. Sci Transl Med. 11:eaat91432019. View Article : Google Scholar | |
|
Loomis KH, Lindsay KE, Zurla C, Bhosle SM, Vanover DA, Blanchard EL, Kirschman JL, Bellamkonda RV and Santangelo PJ: In Vitro Transcribed mRNA vaccines with programmable stimulation of innate immunity. Bioconjug Chem. 29:3072–3083. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Jansen Y, Kruse V, Corthals J, Schats K, van Dam PJ, Seremet T, Heirman C, Brochez L, Kockx M, Thielemans K and Neyns B: A randomized controlled phase II clinical trial on mRNA electroporated autologous monocyte-derived dendritic cells (TriMixDC-MEL) as adjuvant treatment for stage III/IV melanoma patients who are disease-free following the resection of macrometastases. Cancer Immunol Immunother. 69:2589–2598. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Schlake T, Thran M, Fiedler K, Heidenreich R, Petsch B and Fotin-Mleczek M: mRNA: A novel avenue to antibody therapy? Mol Ther. 27:773–784. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Thran M, Mukherjee J, Pönisch M, Fiedler K, Thess A, Mui BL, Hope MJ, Tam YK, Horscroft N, Heidenreich R, et al: mRNA mediates passive vaccination against infectious agents, toxins, and tumors. EMBO Mol Med. 9:1434–1447. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Dahlen E, Veitonmaki N and Norlen P: Bispecific antibodies in cancer immunotherapy. Ther Adv Vaccines Immunother. 6:3–17. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Thomas A, Teicher BA and Hassan A: Antibody-drug conjugates for cancer therapy. Lancet Oncol. 17:e254–e262. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Page DB, Postow MA, Callahan MK, Allison JP and Wolchok JD: Immune modulation in cancer with antibodies. Annu Rev Med. 65:185–202. 2014. View Article : Google Scholar | |
|
Hudis CA: Trastuzumab-mechanism of action and use in clinical practice. N Engl J Med. 357:39–51. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Diamantis N and Banerji U: Antibody-drug conjugates-an emerging class of cancer treatment. Br J Cancer. 114:362–367. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
de Miguel D, Lemke J, Anel A, Walczak H and MartinezLostao L: Onto better TRAILs for cancer treatment. Cell Death Differ. 23:733–747. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Salles G, Barrett M, Foà R, Maurer J, O'Brien S, Valente N, Wenger M and Maloney DG: Rituximab in B-Cell Hematologic Malignancies: A review of 20 years of clinical experience. Adv Ther. 34:2232–2273. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Van Hoecke L, Verbeke R, Dewitte H, Lentacker I, Vermaelen K, Breckpot K and Van Lint S: mRNA in cancer immunotherapy: Beyond a source of antigen. Mol Cancer. 20:482021. View Article : Google Scholar : PubMed/NCBI | |
|
Rybakova Y, Kowalski PS, Huang Y, Gonzalez JT, Heartlein MW, DeRosa F, Delcassian D and Anderson DG: mRNA Delivery for Therapeutic Anti-HER2 antibody expression in vivo. Mol Ther. 27:1415–1423. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Ballesteros-Briones MC, Martisova E, Casales E, Silva-Pilipich N, Buñuales M, Galindo J, Mancheño U, Gorraiz M, Lasarte JJ, Kochan G, et al: Short-Term Local Expression of a PD-L1 Blocking Antibody from a Self-Replicating RNA Vector induces potent antitumor responses. Mol Ther. 27:1892–1905. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Nisonoff A, Wissler FC and Lipman LN: Properties of the major component of a peptic digest of rabbit antibody. Science. 132:1770–1771. 1960. View Article : Google Scholar : PubMed/NCBI | |
|
Wang Y, Tiruthani K, Li S, Hu M, Zhong G, Tang Y, Roy S, Zhang L, Tan J, Liao C and Liu R: mRNA delivery of a bispecific single-domain antibody to polarize tumor-associated macrophages and synergize immunotherapy against liver malignancies. Adv Mater. 33:e20076032021. View Article : Google Scholar : PubMed/NCBI | |
|
Fudenberg HH, Drews G and Nisonoff A: Serologic demonstration of dual specificity of rabbit bivalent hybrid antibody. J Exp Med. 119:151–166. 1964. View Article : Google Scholar : PubMed/NCBI | |
|
Garber K: Bispecific antibodies rise again. Nat Rev Drug Discov. 13:799–801. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Wu L, Wang W, Tian J, Qi C, Cai Z, Yan W, Xuan S and Shang A: Engineered mRNA-expressed bispecific antibody prevent intestinal cancer via lipid nanoparticle delivery. Bioengineered. 12:12383–12393. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Zhao Y, Moon E, Carpenito C, Paulos CM, Liu X, Brennan AL, Chew A, Carroll RG, Scholler J, Levine BL, et al: Multiple injections of electroporated autologous T cells expressing a chimeric antigen receptor mediate regression of human disseminated tumor. Cancer Res. 70:9053–9061. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Rabinovich PM, Komarovskaya ME, Wrzesinski SH, Alderman JL, Budak-Alpdogan T, Karpikov A, Guo H, Flavell RA, Cheung NK, Weissman SM and Bahceci E: Chimeric receptor mRNA transfection as a tool to generate antineoplastic lymphocytes. Hum Gene Ther. 20:51–61. 2009. View Article : Google Scholar | |
|
Morgan RA, Yang JC, Kitano M, Dudley ME, Laurencot CM and Rosenberg SA: Case report of a serious adverse event following the administration of T cells transduced with a chimeric antigen receptor recognizing ERBB2. Mol Ther. 18:843–851. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Brentjens R, Yeh R, Bernal Y, Riviere I and Sadelain M: Treatment of chronic lymphocytic leukemia with genetically targeted autologous T cells: Case report of an unforeseen adverse event in a phase I clinical trial. Mol Ther. 18:666–668. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Miliotou AN and Papadopoulou LC: CAR T-cell Therapy: A new era in cancer immunotherapy. Curr Pharm Biotechnol. 19:5–18. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Fiedler K, Lazzaro S, Lutz J, Rauch S and Heidenreich R: mRNA cancer vaccines. Recent Results Cancer Res. 209:61–85. 2016. View Article : Google Scholar | |
|
Soundara Rajan T, Gugliandolo A, Bramanti P and Mazzon E: In Vitro-Transcribed mRNA Chimeric Antigen Receptor T Cell (IVT mRNA CAR T) therapy in hematologic and solid tumor management: A preclinical update. Int J Mol Sci. 21:65142020. View Article : Google Scholar : PubMed/NCBI | |
|
Zhao Y, Zheng Z, Cohen CJ, Gattinoni L, Palmer DC, Restifo NP, Rosenberg SA and Morgan RA: High-efficiency transfection of primary human and mouse T lymphocytes using RNA electroporation. Mol Ther. 13:151–159. 2006. View Article : Google Scholar | |
|
Yoon SH, Lee JM, Woo SJ, Park MJ, Park JS, Kim HS, Park MY, Sohn HJ and Kim TG: Transfer of Her-2/neu specificity into cytokine-induced killer (CIK) cells with RNA encoding chimeric immune receptor (CIR). J Clin Immunol. 29:806–814. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Schaft N, Dörrie J, Müller I, Beck V, Baumann S, Schunder T, Kämpgen E and Schuler G: A new way to generate cytolytic tumor-specific T cells: Electroporation of RNA coding for a T cell receptor into T lymphocytes. Cancer Immunol Immunother. 55:1132–1141. 2006. View Article : Google Scholar | |
|
Boissel L, Betancur M, Wels WS, Tuncer H and Klingemann H: Transfection with mRNA for CD19 specific chimeric antigen receptor restores NK cell mediated killing of CLL cells. Leuk Res. 33:1255–1259. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Kenderian SS, Ruella M, Shestova O, Klichinsky M, Aikawa V, Morrissette JJ, Scholler J, Song D, Porter DL, Carroll M, et al: CD33-specific chimeric antigen receptor T cells exhibit potent preclinical activity against human acute myeloid leukemia. Leukemia. 29:1637–1647. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Birkholz K, Hombach A, Krug C, Reuter S, Kershaw M, Kämpgen E, Schuler G, Abken H, Schaft N and Dörrie J: Transfer of mRNA encoding recombinant immunoreceptors reprograms CD4+ and CD8+ T cells for use in the adoptive immunotherapy of cancer. Gene Ther. 16:596–604. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Barrett DM, Zhao Y, Liu X, Jiang S, Carpenito C, Kalos M, Carroll RG, June CH and Grupp SA: Treatment of advanced leukemia in mice with mRNA engineered T cells. Hum Gene Ther. 22:1575–1586. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Ang WX, Li Z, Chi Z, Du SH, Chen C, Tay JC, Toh HC, Connolly JE, Xu XH and Wang S: Intraperitoneal immunotherapy with T cells stably and transiently expressing anti-EpCAM CAR in xenograft models of peritoneal carcinomatosis. Oncotarget. 8:13545–13559. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Almasbak H, Rian E, Hoel HJ, Pulè M, Wälchli S, Kvalheim G, Gaudernack G and Rasmussen AM: Transiently redirected T cells for adoptive transfer. Cytotherapy. 13:629–640. 2011. View Article : Google Scholar | |
|
Miliotou AN and Papadopoulou LC: In Vitro-Transcribed (IVT)-mRNA CAR Therapy Development. Methods Mol Biol. 2086:87–117. 2020. View Article : Google Scholar | |
|
Foster JB, Choudhari N, Perazzelli J, Storm J, Hofmann TJ, Jain P, Storm PB, Pardi N, Weissman D, Waanders AJ, et al: Purification of mRNA Encoding Chimeric Antigen Receptor Is Critical for Generation of a Robust T-Cell Response. Hum Gene Ther. 30:168–178. 2019. View Article : Google Scholar : | |
|
Jetani H, Navarro-Bailón A, Maucher M, Frenz S, Verbruggen C, Yeguas A, Vidriales MB, González M, Rial Saborido J, Kraus S, et al: Siglec-6 is a novel target for CAR T-cell therapy in acute myeloid leukemia. Blood. 138:1830–1842. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Lin L, Cho SF, Xing L, Wen K, Li Y, Yu T, Hsieh PA, Chen H, Kurtoglu M, Zhang Y, et al: Preclinical evaluation of CD8+ anti-BCMA mRNA CAR T cells for treatment of multiple myeloma. Leukemia. 35:752–763. 2021. View Article : Google Scholar | |
|
Bontkes HJ, Kramer D, Ruizendaal JJ, Meijer CJ and Hooijberg E: Tumor associated antigen and interleukin-12 mRNA transfected dendritic cells enhance effector function of natural killer cells and antigen specific T-cells. Clin Immunol. 127:375–384. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Wilgenhof S, Van Nuffel AMT, Benteyn D, Corthals J, Aerts C, Heirman C, Van Riet I, Bonehill A, Thielemans K and Neyns B: A phase IB study on intravenous synthetic mRNA electroporated dendritic cell immunotherapy in pretreated advanced melanoma patients. Ann Oncol. 24:2686–2693. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Figlin RA, Tannir NM, Uzzo RG, Tykodi SS, Chen DYT, Master V, Kapoor A, Vaena D, Lowrance W, Bratslavsky G, et al: Results of the ADAPT Phase 3 Study of Rocapuldencel-T in combination with sunitinib as first-line therapy in patients with metastatic renal cell carcinoma. Clin Cancer Res. 26:2327–2336. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Van Hoecke L, Van Lint S, Roose K, Van Parys A, Vandenabeele P, Grooten J, Tavernier J, De Koker S and Saelens X: Treatment with mRNA coding for the necroptosis mediator MLKL induces antitumor immunity directed against neo-epitopes. Nat Commun. 9:34172018. View Article : Google Scholar : PubMed/NCBI | |
|
Trepotec Z, Lichtenegger E, Plank C, Aneja MK and Rudolph C: Delivery of mRNA therapeutics for the treatment of hepatic diseases. Mol Ther. 27:794–802. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Sahu I, Haque AKMA, Weidensee B, Weinmann P and Kormann MSD: Recent Developments in mRNA-Based protein supplementation therapy to target lung diseases. Mol Ther. 27:803–823. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Patel S, Ryals RC, Weller KK, Pennesi ME and Sahay G: Lipid nanoparticles for delivery of messenger RNA to the back of the eye. J Control Release. 303:91–100. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Magadum A, Kaur K and Zangi L: mRNA-Based protein replacement therapy for the heart. Mol Ther. 27:785–793. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Lescan M, Perl RM, Golombek S, Pilz M, Hann L, Yasmin M, Behring A, Keller T, Nolte A, Gruhn F, et al: De Novo Synthesis of elastin by exogenous delivery of synthetic modified mRNA into Skin and Elastin-Deficient Cells. Mol Ther Nucleic Acids. 11:475–484. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Baba M, Itaka K, Kondo K, Yamasoba T and Kataoka K: Treatment of neurological disorders by introducing mRNA in vivo using polyplex nanomicelles. J Control Release. 201:41–48. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Kormann MS, Hasenpusch G, Aneja MK, Nica G, Flemmer AW, Herber-Jonat S, Huppmann M, Mays LE, Illenyi M, Schams A, et al: Expression of therapeutic proteins after delivery of chemically modified mRNA in mice. Nat Biotechnol. 29:154–157. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Van Hoecke L and Roose K: How mRNA therapeutics are entering the monoclonal antibody field. J Transl Med. 17:542019. View Article : Google Scholar : PubMed/NCBI | |
|
Raab M, Kostova I, Peña-Llopis S, Fietz D, Kressin M, Aberoumandi SM, Ullrich E, Becker S, Sanhaji M and Strebhardt K: Rescue of p53 functions by in vitro-transcribed mRNA impedes the growth of high-grade serous ovarian cancer. Cancer Commun (Lond). 44:101–126. 2024. View Article : Google Scholar | |
|
Yin H, Kauffman KJ and Anderson DG: Delivery technologies for genome editing. Nat Rev Drug Discov. 16:387–399. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Cox DB, Platt RJ and Zhang F: Therapeutic genome editing: Prospects and challenges. Nat Med. 21:121–131. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Wang SW, Gao C, Zheng YM, Yi L, Lu JC, Huang XY, Cai JB, Zhang PF, Cui YH and Ke AW: Current applications and future perspective of CRISPR/Cas9 gene editing in cancer. Mol Cancer. 21:572022. View Article : Google Scholar : PubMed/NCBI | |
|
Ling K, Dou Y, Yang N, Deng L, Wang Y, Li Y, Yang L, Chen C, Jiang L, Deng Q, et al: Genome editing mRNA nanotherapies inhibit cervical cancer progression and regulate the immunosuppressive microenvironment for adoptive T-cell therapy. J Control Release. 360:496–513. 2023. View Article : Google Scholar : PubMed/NCBI | |
|
Jo JI, Gao JQ and Tabata Y: Biomaterial-based delivery systems of nucleic acid for regenerative research and regenerative therapy. Regen Ther. 11:123–130. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Kwon H, Kim M, Seo Y, Moon YS, Lee HJ, Lee K and Lee H: Emergence of synthetic mRNA: In vitro synthesis of mRNA and its applications in regenerative medicine. Biomaterials. 156:172–193. 2018. View Article : Google Scholar | |
|
Lin Z, Wu Y, Xu Y, Li G, Li Z and Liu T: Mesenchymal stem cell-derived exosomes in cancer therapy resistance: Recent advances and therapeutic potential. Mol Cancer. 21:1792022. View Article : Google Scholar : PubMed/NCBI | |
|
Ramos da Silva J, Bitencourt Rodrigues K, Formoso Pelegrin G, Silva Sales N, Muramatsu H, de Oliveira Silva M, Porchia BFMM, Moreno ACR, Aps LRMM, Venceslau-Carvalho AA, et al: Single immunizations of self-amplifying or non-replicating mRNA-LNP vaccines control HPV-associated tumors in mice. Sci Transl Med. 15:eabn34642023. View Article : Google Scholar : PubMed/NCBI |









