



Taurine alleviates lipopolysaccharide‑induced liver injury by anti‑inflammation and antioxidants in rats
- Authors:
- Published online on: August 31, 2017 https://doi.org/10.3892/mmr.2017.7414
- Pages: 6512-6517
-
Copyright: © Liu et al. This is an open access article distributed under the terms of Creative Commons Attribution License.
Abstract
Introduction
Hepatic disorder has become a major disease endangering human health and exhibits a high morbidity and fatality rate (1). As the mechanisms of severe hepatic failure are poorly understood, there remains a lack of effective treatment. An increasing number of studies have suggested that the production of inflammatory cytokines serves important roles in various types of liver damage (2,3). Lipopolysaccharide (LPS), a component of Gram-negative bacteria, can activate liver macrophages, which produce inflammatory cytokines including tumor necrosis factor-α (TNF-α), interleukin (IL)-1β and IL-6 (4–6). In addition, LPS possesses pro-oxidative action via the induction of excessive production of reactive oxygen species (ROS). ROS are the major cause of damage to macromolecules, including protein and DNA, and to the cell membrane, which leads to mitochondrial dysfunction (7,8). A further study demonstrated that ROS are involved in modulation of the inflammatory response (9). Various types of liver damage, including ischemia-reperfusion and liver cancer, are associated with LPS (10,11). Therefore, LPS-induced liver injury is used as an animal model of liver disorder.
Taurine, a sulfur-containing β-amino acid, is a metabolic product of L-cysteine and is abundant in a number of mammalian tissues. Taurine is not involved in the synthesis of protein; however, considerable evidence has demonstrated that taurine serves a number of vital roles in physiological processes, including regulation of calcium concentration (12), stabilization of the cell membrane (13), regulation of blood pressure and protection of endothelial cells (14). The antioxidant properties of taurine have been confirmed by a number of results, although taurine itself is not able to scavenge ROS. Taurine exerts its antioxidant action by inhibiting the production of ROS, which result from the increasing activities of antioxidases (15). Certain studies have indicated that taurine protects cells against oxidative stress (16,17). Taurine is changed into taurine chloramine (TauCl) in vivo, which inhibits secretion of pro-inflammatory cytokines including IL-6, IL-1β, TNF-α and IL-8 (18,19). It is suggested that taurine is a potent anti-inflammatory factor.
The present study examined the beneficial effects of taurine on LPS-induced liver injury in rats. The results suggested that administration of taurine may be beneficial for patients with hepatopathy.
Materials and methods
Animals and experimental design
A total of 30 healthy male Sprague-Dawley rats (weighing 280±20 g) were obtained from the Animal Center at West Anhui Health Vocational College and housed in a standard facility at 22°C and 50–70% humidity with a 12-h light/dark cycle. Experimental rats received a standard pellet diet and water ad libitum. The study was approved by the Ethics Committee of West Anhui Health Vocational College (Lu'an, China). After a week, the animals were randomly divided into three groups (n=10 per group): i) Normal saline group (NS), ii) LPS control group (LPS) and iii) taurine + LPS group (TL). Rats from NS and LPS were treated with sterile saline by intravenous injection and animals from TL were intravenously injected with taurine (100 mg/kg body weight, dissolved in sterile saline; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). After 4 h, rats from LPS and TL groups were intraperitoneally injected with LPS (10 mg/kg body weight, dissolved in sterile saline; Sigma-Aldrich; Merck KGaA) and NS rats received sterile saline. At 6 h following administration of LPS, the animals were anesthetized with sodium pentobarbital (50 mg/kg, i.p.; Sigma-Aldrich; Merck KGaA) to collect blood samples prior to animal sacrifice. Fasting blood samples were collected by artery catheterization for biochemical analyses and liver tissues were obtained for histological analyses.
Determination of liver function
Fasting blood samples were centrifuged at 3,000 × g to separate the serum. Markers of liver function, serum aspartate transaminase (AST) and alanine transaminase (ALT) were determined using an enzymatic colorimetric method (Diagnostica Stago S.A.S., Paris, France) according to the manufacturer's protocols and analyzed by a semi-automatic analyzer, with the results being expressed as U/l.
Measurement of inflammatory cytokines
Serum levels of TNF-α, procalcitonin (PCT) and IL-6 were detected with rat TNF-α (cat. no. ELS-2855-1), PCT (cat. no. ELS-3485-1) and IL-6 (cat. no. ELS-2866-1) specific ELISA kits (Hefei Bomei Biotechnology Co., Ltd., Hefei, China) according to manufacturer's protocol. The levels of TNF-α and IL-6 were expressed as ng/l.
Estimation of antioxidant effects
To estimate changes in antioxidant effects, the activity of the antioxidase, superoxide dismutase (SOD) was determined using xanthine oxidase methods, and the content of lipid peroxidation production, malonaldehyde (MDA), was measured using thiobarbituric acid methods, according to the manufacturer's protocol (both from Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Histological analysis
Liver tissues were harvested and fixed in 4% phosphate-buffered formalin for pathological analysis. Fixed tissues were dehydrated in ethanol, embedded in paraffin and 5 µm sections were cut. After drying overnight, sections were dewaxed, rehydrated and stained with hematoxylin and eosin (H&E) for histomorphological observation under a light microscope.
Western blot analysis
Liver tissues (0.2 g) were harvested, lysed and homogenized in 2 ml lysis buffer with 10 mM Tris-buffered saline, 1 mM EDTA, 1 mM EGTA, 2 mM PMSF and 1% Triton X-100 (v/v) for 20 min. Lysates were centrifuged at 13,000 × g for 15 min at 4°C. Protein concentration was measured using a Quick Start™ Bradford protein assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Denatured proteins in supernatants were separated by 10% SDS-PAGE and transferred onto nitrocellulose membranes. The membranes were blocked with 5% non-fat milk in TBS with Tween-20 (10 mM Tris-HCl, 150 mM NaCl and 1% Tween-20) for 2 h. The membranes were subsequently incubated with primary polyclonal antibodies against β-actin (1:1,000), heme oxygenase-1 (HO-1; 1:1,000), cyclooxygenase-2 (COX-2), nuclear factor (NF)-κB, phosphorylated (p)-NF-κB, extracellular signal-regulated kinase (ERK) and p-ERK1/2 (Bio Basic Inc., Markham, ON, Canada) overnight at 4°C. Following an extensive wash with TBST, the membranes were incubated with a horseradish peroxidase-conjugated goat anti-rabbit secondary antibody (1:10,000; cat. no. A9169; Sigma-Aldrich; Merck KGaA) for 2 h at room temperature. The membranes were washed three times and visualized with 3,3′-diaminobenzidine (Bio Basic Inc.).
Statistical analysis
All values are expressed as mean ± standard deviation. The data were analyzed using SPSS version 16.0 (SPSS, Inc., Chicago, IL, USA). Statistical difference was determined by Tukey's test for unpaired data or one-way analysis of variance with least significant difference-t and/or Tamhane's T2 post hoc tests for multiple comparisons. P<0.05 was considered statistically to indicate a statistically significant difference.
Results
Ameliorative effects of taurine on liver damage
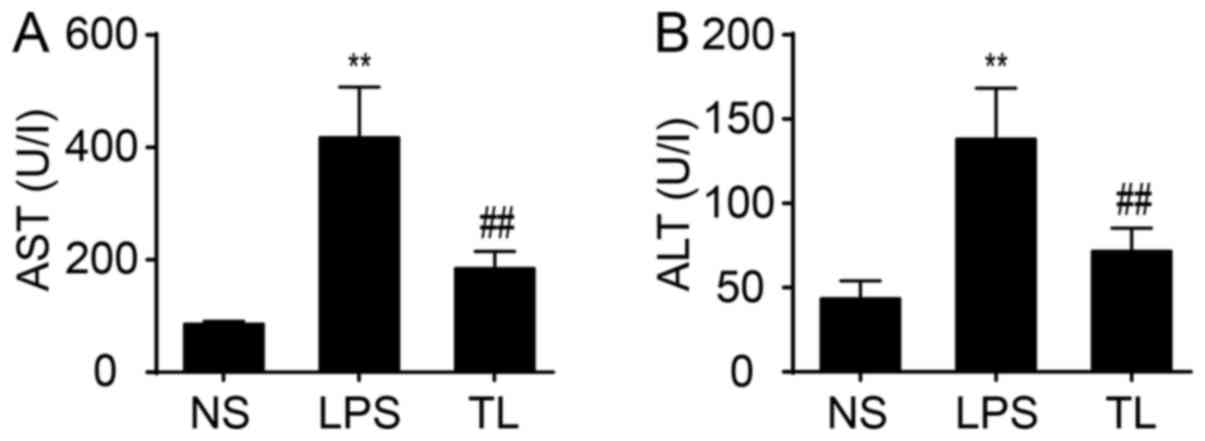
To examine liver function, serum AST and ALT transaminases were determined. The result demonstrated a significant increase in activity of AST and ALT in LPS rats compared with NS rats (P<0.01; Fig. 1). Administration of taurine reduced the increase in activity of AST and ALT (P<0.01; Fig. 1).
Change of antioxidant effects
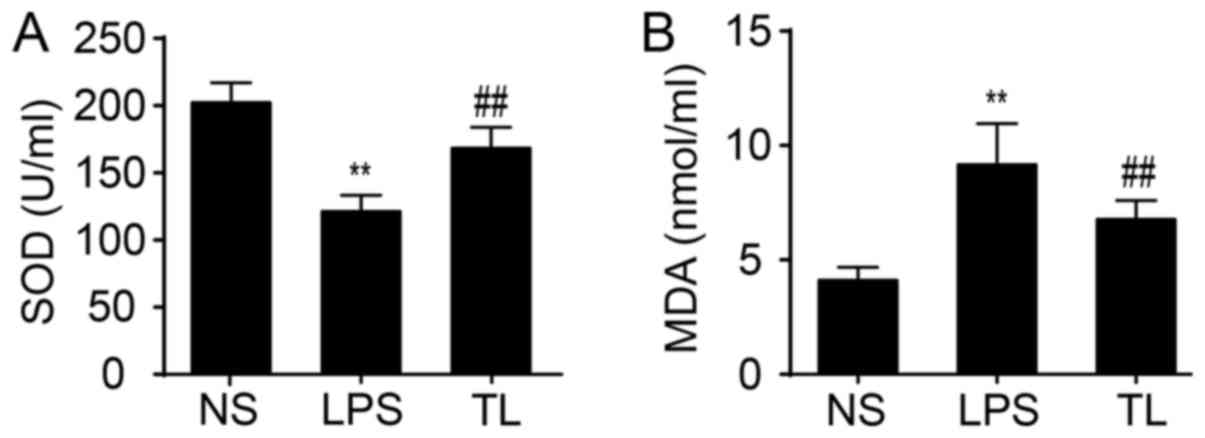
The activity of serum SOD, an antioxidase, was reduced and the content of serum MDA, a product of lipid peroxidation, was increased in LPS-treated rats compared with NS rats (P<0.01; Fig. 2). Taurine significantly enhanced SOD activity and decreased the concentration of MDA (P<0.01; Fig. 2).
Effects of taurine on pro-inflammatory cytokines
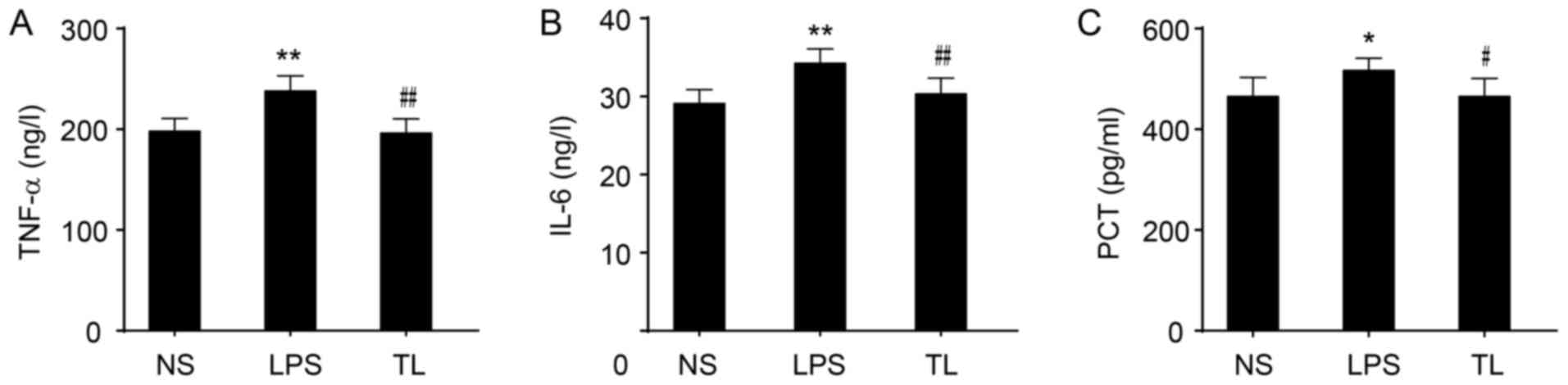
The levels of TNF-α and IL-6 in serum were significantly increased in LPS rats (P<0.01; Fig. 3A and B) and an increased level of PCT was determined (P<0.05; Fig. 3C). Taurine treatment prior to LPS significantly reduced the levels of TNF-α and IL-6 (P<0.01; Fig. 3A and B) and decreased the concentration of PCT (P<0.05; Fig. 3C).
Effects of taurine on hepatic histopathology
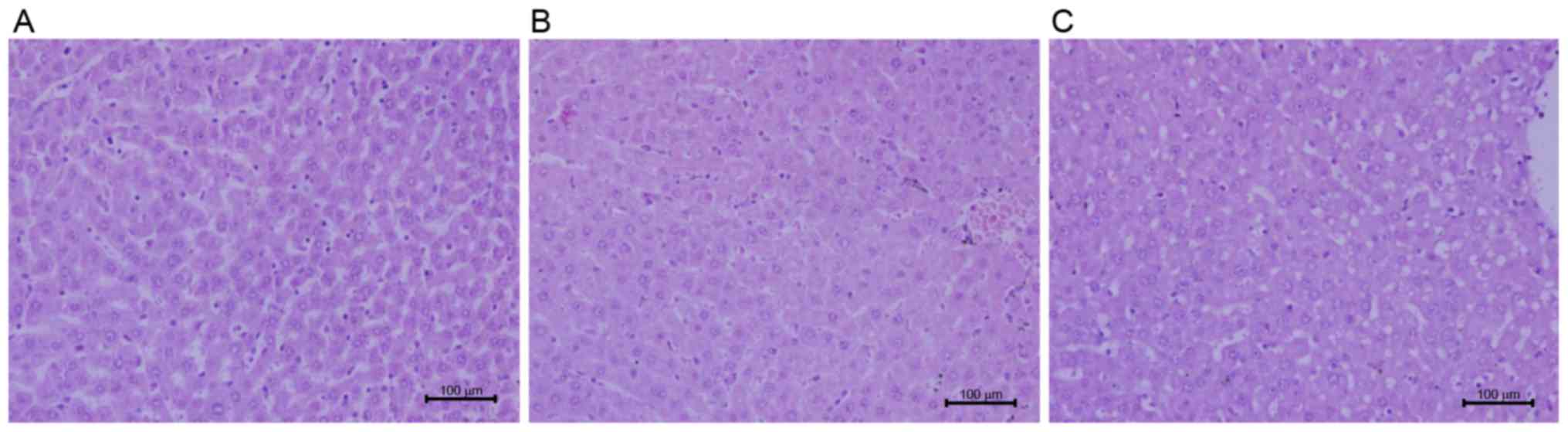
Liver sections stained with H&E were observed under a light microscope for hepatic morphology. Recruitment of inflammatory cells and release of inflammatory factors in liver are involved in liver injury (20). Exposure to LPS resulted in an increase in the infiltration of inflammatory cells and hepatocyte edema (Fig. 4). Administration of taurine prior to LPS attenuated congestion in liver tissues, abated the number of the infiltration of inflammatory cells and intact lobular structure was observed (Fig. 4).
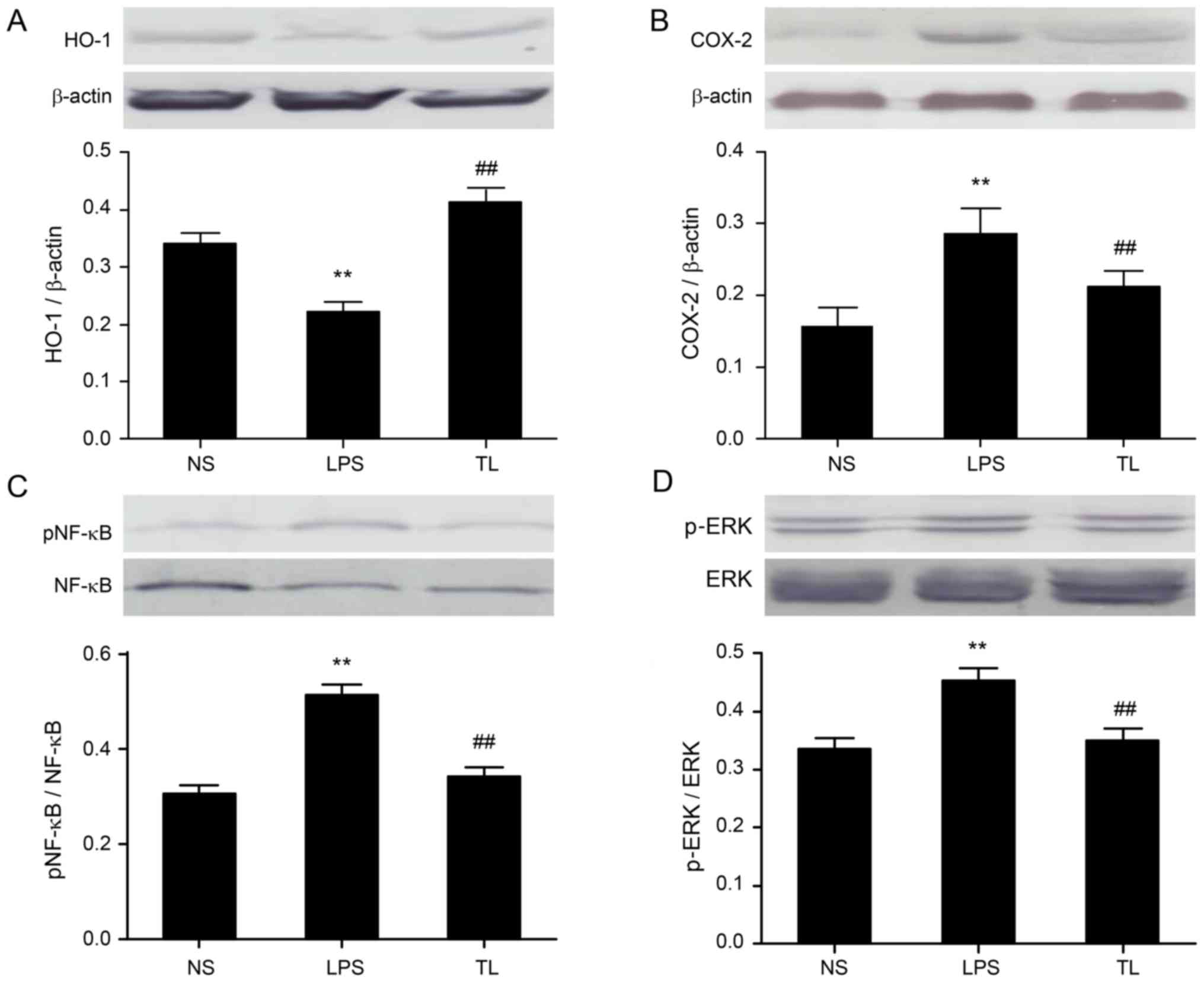
Effects of taurine on liver ERK1/2, COX-2, HO-1 and NF-κB protein expression
Antioxidant alterations were examined; taurine pretreatment elevated HO-1 protein expression (Fig. 5A). Furthermore, the protein expression levels of COX-2 (Fig. 5B), NF-κB (Fig. 5C) and p-ERK (Fig. 5D) were investigated to evaluate inflammation signaling. Taurine pretreatment reduced the protein expression of NF-κB, COX-2 and p-ERK.
Discussion
It is well-known that activation of macrophages and release of inflammatory cytokines serve important roles in organ damage, including acute and chronic liver injury (21,22). The present study aimed to investigate effects of taurine pretreatment on LPS-induced liver injury. The results demonstrated that taurine pretreatment by intravenous injection reduced the activity of plasma AST and ALT, and decreased the level of serum inflammatory cytokines including TNF-α, IL-6 and MDA. Serum SOD activity and HO-1 protein expression in liver was significantly increased in taurine-pretreated rats, while COX-2, pNF-κB (E) and p-ERK protein expression levels in liver were reduced. In addition, taurine pretreatment alleviated the infiltration of inflammatory cells in liver tissues and hepatic congestion. The present study suggested that taurine pretreatment protected the liver against LPS-induced injury.
LPS-induced tissue injury results from an increase in the release of cytokines, oxidative stress and impairment of mitochondrial function (23). LPS induces excessive release of pro-inflammatory cytokines including TNF-α and IL-6 and the production of ROS by binding with Toll-like receptor 4 on the surface of Küpffer cells (24,25), stimulating the apoptosis of hepatic cells and necrosis (26).
Oxidative stress results from excessive generation of ROS and/or deletion of antioxidants including reduced activities of the antioxidant enzymes, imbalance of glutathione redox status (27) and increased products of lipid peroxidation (28), which damages cells via macromolecules and mitochondrial dysfunction, further harming various tissues including the liver (7,29). Release of ROS is a mechanism of LPS-induced hepatic injury, therefore reduction of ROS signaling relieves such damage (30). Studies have indicated that treatment with antioxidant and anti-inflammatory agents is beneficial in LPS-induced hepatic injury (31,32), and that taurine can reduce oxidative stress (33,34) and relieve tissue injuries by its antioxidative properties (35,36). The results of the present study indicated that administration of LPS resulted increased ALT and AST activities in serum, considered as markers of liver injury. Taurine pretreatment reduced the increases of ALT and AST, and decreased the concentration of MDA, a marker of lipid peroxidation; it also elevated the activity of SOD and the protein expression of antioxidant enzyme HO-1.
Inflammatory response serves an important role in various liver disorders. LPS initiates inflammation by recruiting neutrophils to the liver and subsequently stimulating the expression of inflammatory factors, including TNF-α, which provoke the release of ROS (37). The results of the present study demonstrated that LPS elevated the levels of TNF-α and IL-6 in serum and expression of COX-2 and NF-κB protein. Taurine pretreatment reduced the expression of pro-inflammatory proteins including COX-2 and NF-κB.
It is reported that taurine is converted into taurine chloramine in vivo and that this reduces the inflammatory response (38). Taurine increases its antioxidative effects by increasing the expression of HO-1 protein (39), which is reported to inhibit expression of COX-2 (40). COX-2 induces the production of prostaglandin, which is involved in inflammation and pain and results in cellular injury (41,42). HO-1, an inducible rate-limiting enzyme, catalyzes heme into equimolar amounts of carbon monoxide (CO), biliverdin and free iron. Induction of HO-1 may protect against oxidative stress-related cell and tissue injury (40,43). Biliverdin has been confirmed to be a potent antioxidant (44). CO, a catalytic product of HO-1, exerts antioxidative, anti-inflammatory and anti-apoptotic effects (45). Increasing evidence has confirmed that the HO/CO signaling exerts a vital role in regulation of anti-inflammation and cytoprotection (46–48).
NF-κB is involved in LPS signaling. Activation of NF-κB by LPS upregulates the expression of COX-2 (49). It is reported that the anti-inflammatory effect of CO is associated with its regulation on transcription factors including NF-κB (50,51). A previous study demonstrated that CO alleviates LPS-induced inflammation by suppression of NF-κB (48). The results suggested that taurine inhibited NF-κB/COX-2 signaling via induction of HO-1. In addition, the mitogen-activated protein kinase (MAPK)/ERK pathway is implicated in LPS-induced inflammation (52) and is an upstream mediator of NF-κB nuclear translocation (53,54). Another study demonstrated that reduced MAPK/ERK1/2 signaling downregulates NF-κB in LPS-activated hepatocytes (54). Recent studies have indicated that increased levels of phosphorylated ERK and c-Jun N-terminal kinases promote COX-2 protein expression (55,56).
In conclusion, the results of the present study indicated that taurine pretreatment protected the liver against LPS-induced injury by increasing its antioxidation and anti-inflammation ability, which were associated with the increased expression of HO-1 protein and reduced expression levels of NF-κB, COX-2 and p-ERK proteins. The findings suggested that taurine reduced NF-κB/COX-2 signaling by activation of HO-1/CO.
Acknowledgements
The present study was supported by the Outstanding Young Talent Fund Key Project of Anhui Province (grant no. 2013SQRL146ZD).
References
|
Mizuhara H, O'Neill E, Seki N, Ogawa T, Kusunoki C, Otsuka K, Satoh S, Niwa M, Senoh H and Fujiwara H: T cell activation-associated hepatic injury: Mediation by tumor necrosis factors and protection by interleukin 6. J Exp Med. 179:1529–1537. 1994. View Article : Google Scholar : PubMed/NCBI | |
|
Gasparini C and Feldmann M: NF-κB as a target for modulating inflammatory responses. Curr Pharm Des. 18:5735–5745. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Josephs MD, Bahjat FR, Fukuzuka K, Ksontini R, Solorzano CC, Edwards CK III, Tannahill CL, MacKay SL, Copeland EM III and Moldawer LL: Lipopolysaccharide and D-galactosamine-induced hepatic injury is mediated by TNF-alpha and not by Fas ligand. Am J Physiol Regul Integr Comp Physiol. 278:R1196–R1201. 2000.PubMed/NCBI | |
|
Enomoto N, Ikejima K, Bradford BU, Rivera CA, Kono H, Goto M, Yamashina S, Schemmer P, Kitamura T, Oide H, et al: Role of Kupffer cells and gut-derived endotoxins in alcoholic liver injury. J Gastroenterol Hepatol. 15 Suppl:D20–D25. 2000. View Article : Google Scholar : PubMed/NCBI | |
|
Uesugi T, Froh M, Arteel GE, Bradford BU, Wheeler MD, Gäbele E, Isayama F and Thurman RG: Role of lipopolysaccharide-binding protein in early alcohol-induced liver injury in mice. J Immunol. 168:2963–2969. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Enomoto N, Schemmer P, Ikejima K, Takei Y, Sato N, Brenner DA and Thurman RG: Long-term alcohol exposure changes sensitivity of rat Kupffer cells to lipopolysaccharide. Alcohol Clin Exp Res. 25:1360–1367. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Cadenas S and Cadenas AM: Fighting the stranger-antioxidant protection against endotoxin toxicity. Toxicology. 180:45–63. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Mallis RJ, Buss JE and Thomas JA: Oxidative modification of H-ras: S-thiolation and S-nitrosylation of reactive cysteines. Biochem J. 355:145–153. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Zuo Y, Xiang B, Yang J, Sun X, Wang Y, Cang H and Yi J: Oxidative modification of caspase-9 facilitates its activation via disulfide-mediated interaction with Apaf-1. Cell Res. 19:449–457. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Colletti LM and Green M: Lung and liver injury following hepatic ischemia/reperfusion in the rat is increased by exogenous lipopolysaccharide which also increases hepatic TNF production in vivo and in vitro. Shock. 16:312–319. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Yu LX, Yan HX, Liu Q, Yang W, Wu HP, Dong W, Tang L, Lin Y, He YQ, Zou SS, et al: Endotoxin accumulation prevents carcinogen-induced apoptosis and promotes liver tumorigenesis in rodents. Hepatology. 52:1322–1333. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Huxtable RJ: Physiological actions of taurine. Physiol Rev. 72:101–163. 1992.PubMed/NCBI | |
|
Pasantes-Morales H, Wright CE and Gaull GE: Taurine protection of lymphoblastoid cells from iron-ascorbate induced damage. Biochem Pharmacol. 34:2205–2207. 1985. View Article : Google Scholar : PubMed/NCBI | |
|
Maia AR, Batista TM, Victorio JA, Clerici SP, Delbin MA, Carneiro EM and Davel AP: Taurine supplementation reduces blood pressure and prevents endothelial dysfunction and oxidative stress in post-weaning protein-restricted rats. PLoS One. 9:e1058512014. View Article : Google Scholar : PubMed/NCBI | |
|
Sun Jang J, Piao S, Cha YN and Kim C: Taurine chloramine activates Nrf2, increases HO-1 expression and protects cells from death caused by hydrogen peroxide. J Clin Biochem Nutr. 45:37–43. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Schaffer SW, Azuma J and Mozaffari M: Role of antioxidant activity of taurine in diabetes. Can J Physiol Pharmacol. 87:91–99. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Erdamar H, Turközkan N, Ekremoğlu M, Kurt Y and Yaman H: The effect of taurine on polymorphonuclear leukocyte functions in endotoxemia. Amino Acids. 33:581–585. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Kontny E, Plebanczyk M, Lisowska B, Olszewska M, Maldyk P and Maslinski W: Comparison of rheumatoid articular adipose and synovial tissue reactivity to proinflammatory stimuli: Contribution to adipocytokine network. Ann Rheum Dis. 71:262–267. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Marcinkiewicz J and Kontny E: Taurine and inflammatory diseases. Amino Acids. 46:7–20. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Ambade A, Catalano D, Lim A and Mandrekar P: Inhibition of heat shock protein (molecular weight 90 kDa) attenuates proinflammatory cytokines and prevents lipopolysaccharide-induced liver injury in mice. Hepatology. 55:1585–1595. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Rossignol DP and Lynn M: TLR4 antagonists for endotoxemia and beyond. Curr Opin Investig Drugs. 6:496–502. 2005.PubMed/NCBI | |
|
Nolan JP: The role of intestinal endotoxin in liver injury: A long and evolving history. Hepatology. 52:1829–1835. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Lowes DA, Webster NR, Murphy MP and Galley HF: Antioxidants that protect mitochondria reduce interleukin-6 and oxidative stress, improve mitochondrial function, and reduce biochemical markers of organ dysfunction in a rat model of acute sepsis. Br J Anaesth. 110:472–480. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Sun S, Zhang H, Xue B, Wu Y, Wang J, Yin Z and Luo L: Protective effect of glutathione against lipopolysaccharide-induced inflammation and mortality in rats. Inflamm Res. 55:504–510. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Ohsaki Y, Shirakawa H, Hiwatashi K, Furukawa Y, Mizutani T and Komai M: Vitamin K suppresses lipopolysaccharide-induced inflammation in the rat. Biosci Biotechnol Biochem. 70:926–932. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Wang Y, Gao LN, Cui YL and Jiang HL: Protective effect of danhong injection on acute hepatic failure induced by lipopolysaccharide and d-galactosamine in mice. Evid Based Complement Alternat Med. 2014:1539022014.PubMed/NCBI | |
|
Davies KJ: Protein damage and degradation by oxygen radicals. I. General aspects. J Biol Chem. 262:9895–9901. 1987.PubMed/NCBI | |
|
Klein T, Neuhaus K, Reutter F and Nüsing RM: Generation of 8-epi-prostaglandin F(2alpha) in isolated rat kidney glomeruli by a radical-independent mechanism. Br J Pharmacol. 133:643–650. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Sewerynek E, Melchiorri D, Chen L and Reiter RJ: Melatonin reduces both basal and bacterial lipopolysaccharide-induced lipid peroxidation in vitro. Free Radic Biol Med. 19:903–909. 1995. View Article : Google Scholar : PubMed/NCBI | |
|
Hsing CH, Lin MC, Choi PC, Huang WC, Kai JI, Tsai CC, Cheng YL, Hsieh CY, Wang CY, Chang YP, et al: Anesthetic propofol reduces endotoxic inflammation by inhibiting reactive oxygen species-regulated Akt/IKKβ/NF-κB signaling. PLoS One. 6:e175982011. View Article : Google Scholar : PubMed/NCBI | |
|
Ajuwon OR, Oguntibeju OO and Marnewick JL: Amelioration of lipopolysaccharide-induced liver injury by aqueous rooibos (Aspalathus linearis) extract via inhibition of pro-inflammatory cytokines and oxidative stress. BMC Complement Altern Med. 14:3922014. View Article : Google Scholar : PubMed/NCBI | |
|
Takata J, Ito S, Karube Y, Nagata Y and Matsushima Y: Water-soluble prodrug of vitamin E for parenteral use and its effect on endotoxin-induced liver toxicity. Biol Pharm Bull. 20:204–209. 1997. View Article : Google Scholar : PubMed/NCBI | |
|
Oliveira MW, Minotto JB, de Oliveira MR, Zanotto-Filho A, Behr GA, Rocha RF, Moreira JC and Klamt F: Scavenging and antioxidant potential of physiological taurine concentrations against different reactive oxygen/nitrogen species. Pharmacol Rep. 62:185–193. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Jeon SH, Lee MY, Rahman MM, Kim SJ, Kim GB, Park SY, Hong CU, Kim SZ, Kim JS and Kang HS: The antioxidant, taurine reduced lipopolysaccharide (LPS)-induced generation of ROS, and activation of MAPKs and Bax in cultured pneumocytes. Pulm Pharmacol Ther. 22:562–566. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Motawi TK, Abd Elgawad HM and Shahin NN: Modulation of indomethacin-induced gastric injury by spermine and taurine in rats. J Biochem Mol Toxicol. 21:280–288. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Shimizu M, Zhao Z, Ishimoto Y and Satsu H: Dietary taurine attenuates dextran sulfate sodium (DSS)-induced experimental colitis in mice. Adv Exp Med Biol. 643:265–271. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
McDonald B, Jenne CN, Zhuo L, Kimata K and Kubes P: Kupffer cells and activation of endothelial TLR4 coordinate neutrophil adhesion within liver sinusoids during endotoxemia. Am J Physiol Gastrointest Liver Physiol. 305:G797–G806. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Weiss SJ, Klein R, Slivka A and Wei M: Chlorination of taurine by human neutrophils. Evidence for hypochlorous acid generation. J Clin Invest. 70:598–607. 1982. View Article : Google Scholar : PubMed/NCBI | |
|
Wang GG, Li W, Lu XH, Zhao X and Xu L: Taurine attenuates oxidative stress and alleviates cardiac failure in type I diabetic rats. Croat Med J. 54:171–179. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Shih RH and Yang CM: Induction of heme oxygenase-1 attenuates lipopolysaccharide-induced cyclooxygenase-2 expression in mouse brain endothelial cells. J Neuroinflammation. 7:862010. View Article : Google Scholar : PubMed/NCBI | |
|
Fletcher JR: Eicosanoids. Critical agents in the physiological process and cellular injury. Arch Surg. 128:1192–1196. 1993. View Article : Google Scholar : PubMed/NCBI | |
|
Williams JA and Shacter E: Regulation of macrophage cytokine production by prostaglandin E2. Distinct roles of cyclooxygenase-1 and −2. J Biol Chem. 272:25693–25699. 1997. View Article : Google Scholar : PubMed/NCBI | |
|
Chen QY, Wang GG, Li W, Jiang YX, Lu XH and Zhou PP: Heme oxygenase-1 promotes delayed wound healing in diabetic rats. J Diabetes Res. 2016:97265032016. View Article : Google Scholar : PubMed/NCBI | |
|
Stocker R, Yamamoto Y, McDonagh AF, Glazer AN and Ames BN: Bilirubin is an antioxidant of possible physiological importance. Science. 235:1043–1046. 1987. View Article : Google Scholar : PubMed/NCBI | |
|
Ryter SW, Alam J and Choi AM: Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol Rev. 86:583–650. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Ryter SW, Otterbein LE, Morse D and Choi AM: Heme oxygenase/carbon monoxide signaling pathways: Regulation and functional significance. Mol Cell Biochem 234–235. 1–263. 2002. | |
|
Wang XM, Kim HP, Nakahira K, Ryter SW and Choi AM: The heme oxygenase-1/carbon monoxide pathway suppresses TLR4 signaling by regulating the interaction of TLR4 with caveolin-1. J Immunol. 182:3809–3818. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Chhikara M, Wang S, Kern SJ, Ferreyra GA, Barb JJ, Munson PJ and Danner RL: Carbon monoxide blocks lipopolysaccharide-induced gene expression by interfering with proximal TLR4 to NF-kappaB signal transduction in human monocytes. PLoS One. 4:e81392009. View Article : Google Scholar : PubMed/NCBI | |
|
Nakao S, Ogata Y, Shimizu-Sasaki E, Yamazaki M, Furuyama S and Sugiya H: Activation of NFkappaB is necessary for IL-1beta-induced cyclooxygenase-2 (COX-2) expression in human gingival fibroblasts. Mol Cell Biochem. 209:113–118. 2000. View Article : Google Scholar : PubMed/NCBI | |
|
Morse D, Pischke SE, Zhou Z, Davis RJ, Flavell RA, Loop T, Otterbein SL, Otterbein LE and Choi AM: Suppression of inflammatory cytokine production by carbon monoxide involves the JNK pathway and AP-1. J Biol Chem. 278:36993–36998. 2003. View Article : Google Scholar : PubMed/NCBI | |
|
Sarady JK, Otterbein SL, Liu F, Otterbein LE and Choi AM: Carbon monoxide modulates endotoxin-induced production of granulocyte macrophage colony-stimulating factor in macrophages. Am J Respir Cell Mol Biol. 27:739–745. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Jang SI, Kim HJ, Kim YJ, Jeong SI and You YO: Tanshinone IIA inhibits LPS-induced NF-kappaB activation in RAW 264.7 cells: Possible involvement of the NIK-IKK, ERK1/2, p38 and JNK pathways. Eur J Pharmacol. 542:1–7. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Chiu WT, Lin YL, Chou CW and Chen RM: Propofol inhibits lipoteichoic acid-induced iNOS gene expression in macrophages possibly through downregulation of toll-like receptor 2-mediated activation of Raf-MEK1/2-ERK1/2-IKK-NFkappaB. Chem Biol Interact. 181:430–439. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Jawan B, Kao YH, Goto S, Pan MC, Lin YC, Hsu LW, Nakano T, Lai CY, Sun CK, Cheng YF, et al: Propofol pretreatment attenuates LPS-induced granulocyte-macrophage colony-stimulating factor production in cultured hepatocytes by suppressing MAPK/ERK activity and NF-kappaB translocation. Toxicol Appl Pharmacol. 229:362–373. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Chen WC, Tseng CK, Chen YH, Lin CK, Hsu SH, Wang SN and Lee JC: HCV NS5A Up-regulates COX-2 expression via IL-8-mediated activation of the ERK/JNK MAPK pathway. PLoS One. 10:e01332642015. View Article : Google Scholar : PubMed/NCBI | |
|
Gu W, Song L, Li XM, Wang D, Guo XJ and Xu WG: Mesenchymal stem cells alleviate airway inflammation and emphysema in COPD through down-regulation of cyclooxygenase-2 via p38 and ERK MAPK pathways. Sci Rep. 5:87332015. View Article : Google Scholar : PubMed/NCBI |









