



The phenotypic reversion of cancer: Experimental evidences on cancer reversibility through epigenetic mechanisms (Review)
- Authors:
- Published online on: January 23, 2024 https://doi.org/10.3892/or.2024.8707
- Article Number: 48
-
Copyright: © Pensotti et al. This is an open access article distributed under the terms of Creative Commons Attribution License.
Abstract
Introduction
Cancer is commonly observed as an irreversible process. Consistent with this interpretation, the current therapies are focused on the elimination of cancer cells by the means of surgery, radiotherapy, chemotherapy/immunotherapy. However, these approaches are not yet decisive for the oncological pathology, thus highlighting either the need for improved eradication treatment or the lack of a general understanding of the cancer process.
In this scenario, the current implicit notion of irreversibility related to cancer should be questioned: Is cancer really irreversible? A significant amount of experimental data has revealed that cancer, under specific conditions, can revert into a benign phenotype. This fact represents a clear paradox under the current gene-based model of cancer according to which the primary cause of cancer is a genetic mutation. Being a genetic mutation irreversible and being cancer considered essentially caused by genetic mutations, so this notion has been directly transferred to cancer. Even though the most updated cancer models take into consideration numerous other factors such as epigenetic, genetic mutations are still implicitly considered hierarchically as the primary cause of cancer.
To improve exploring this issue the literature related to experimental evidence regarding cancer reversibility was selected and analyzed. The literature was reviewed in chronological order, and this work was organized by classifying the different experimental models in order to improve grasping of the relevant issues coming from experimental data. The most relevant studies were selected in consideration of the number of citations received or considering the fact that they were the first experiment of such kind ever performed. For the current review, PubMed (https://pubmed.ncbi.nlm.nih.gov/) and Google scholar (https://scholar.google.com/) were mainly used. The process of tumor reversion has been differently named and described, thus different key words were used including ‘tumor regression’, ‘tumor reprogramming’ and ‘tumor differentiation’. Only studies related to the epigenetic-induced tumor reversion and not gene editing were considered. In fact, it was considered that the concept of tumor reversion is related to physiological complex processes that override genetic mutations. In fact, one of the issues related to tumor reversion processes is that it represents a ‘paradox’ within the somatic mutation theory (SMT) and therefore pushes theoretical biologists to reconsider the role of the genes in cancer research. A subsequent analysis of the literature allowed the authors to discern studies that describe real tumor reversion processes. Furthermore, the bibliography of each study was analyzed in order to collect the most relevant literature on tumor reversion. Once new authors that work on this subject were identified through bibliographic analysis, further research was performed using the name of the author.
At the end of this work, the implication of these data were analyzed in consideration of the current model of cancer and of the current structure of the ‘cancer research systems’ understood as the intertwining of science, medicine, industry, finance and law.
Embryonal rest theory of cancer
Originally, the very first concept of ‘metastasis’ implicitly considered the possibility that cancer might spontaneously regress. The word ‘metastasis’ was introduced in 1829 by the French gynecologist, Joseph Claude Anhelme Récamier, after a clinical observation on a patient with breast cancer. After the patient's death, the autopsy revealed the complete disappearance of the breast cancer mass and the presence of a tumor mass localized in the right lobe of the brain (1). Commenting on this clinical case, Récamier introduced the term metastasis for the first time using these words: ‘La résolution spontanée d'un engorgement carcinomateux, suivie d'un autre engorgement de même nature, peut conduire à admettre des métastases cancéreuses’ [the spontaneous resolution of a carcinomatous engorgement, followed by another engorgement of the same nature, can lead to admit that cancerous metastases had occurred] (1).
Moreover, Recamier noticed-together with his pathologist colleague, Jean Lobstein-several histological similarities between the samples of tumor and embryonic tissue. They thus presented the hypothesis that cancer could originate from a residual of embryonic cells still present within the adult organism (2,3). The German anatomist and physiologist, Johannes Müller, proposed to associate embryogenesis and carcinogenesis. He described tumors as the uncontrolled continuation of embryonic developmental processes (4).
A further endorsement of this hypothesis arised in 1855 from the German pathologist, Rudolph Virchow. He confirmed the observations of Récamier and Lobstein at the cellular level and specified that tumor and embryonic cells also share several functional and structural characteristics (5). This led him to hypothesize that cancer could originate directly from embryonic-like cells (6). Such a thesis was later developed and structured by his student, Julius Cohnheim, together with Francesco Durante, who introduced the ‘embryonal rest theory of cancer’. This theory states that adult tissues contain residues of embryonic cells which, under certain conditions, can reactivate and give rise to tumor masses (7,8). Such a model considered cancer as reversible, at least in theory. In fact, embryonic cells can differentiate into normal somatic cells. Therefore, cancer cells deriving from embryonic stem cells should also be able to transform themselves into benign differentiated tissue.
In addition, Max Wilms, a German pathologist and surgeon, indirectly supported the association between embryogenesis and carcinogenesis when, in 1899, he observed a kidney tumor populated by embryonic cells in an 8-year-old boy (9). An attempt to explain the activation mechanisms of residual embryonic cells in adults was made by Hugo Ribbert between the late 1800s and the early 1900s. He advanced the hypothesis that the tissue microenvironment exerts a sort of ‘tension’ on embryonic cells, which is capable of keeping them dormant. When this tension is lost, then the uncontrolled processes of carcinogenesis start (10,11).
These clinical and histological observations converge toward the idea that tumor cells and embryonic cells share some fundamental characteristics. Later on, and following this hypothesis, it has been theoretically possible to consider cancer cells as ‘developmental processes gone awry’ (12). This clearly implies a new strategy for cancer treatments, that is, a re-differentiation approach that implies a modulation of phenotypic expression.
The embryonal theory of cancer allowed scientists to consider the hypothesis of the reversibility of tumors. However, cancer models began to structure according to the SMT after the 1950s. SMT considers cancer an ‘irreversible’ process because it considers gene mutations that are irreversible, as the primary cause of cancer.
Teratoma as a model for tumor reversion research
The cells of teratomas, that is, a type of germ cell tumor that may contain several different types of tissue, are the most similar to embryonic cells. Accordingly, they represent an enlightening link between tumors and embryonic cells. Teratomas are composed of a heterogeneous series of cells from differentiated tissues-each of which represent primary germ layers, to which are added embryonic tumor cells. Ovarian teratomas, for example, in the early stages of development are composed of a fairly homogeneous cell population. This turns into a series of differentiated cells as the teratoma progresses. In some cases, it gives rise to completely differentiated structures such as teeth and hair. Hence, the term teratoma, from the Greek ‘τέρας’ (téras), which means monster. In fact, teratomas appeared monstrous precisely due to the presence of differentiated structures within shapeless masses, almost as if a new living being with ‘monstrous’ characteristics were trying to emerge from these tumor masses. The fact that portions of differentiated, non-malignant, tissues emerged from cancer cells suggests that it was possible to transform cancer cells into normal cells.
The first clinical confirmation of this hypothesis dates back to 1907 when the Swiss pathologist, Max Askanazy, studied an ovarian teratoma in the initial stage. He observed and described the spontaneous regression of the tumor mass where the teratoma cells differentiated and gave rise to normal tissue (13). Teratomas are rare tumors and, therefore, difficult to be systematically studied. The contribution by Leroy Stevens and Clarence Little was therefore important: They succeeded in creating an innate strain of mice that was highly prone to develop teratomas (14). This strain, called 129/SvJ, gave impulse to the study of teratomas and allowed for increasingly refined experimental designs.
It is precisely from this animal model that, in 1959, Barry Pierce first observed the spontaneous differentiation of tumor embryonic cells from testicular teratocarcinomas (15). He isolated some embryonic carcinoma cells from the differentiated tissues of murine teratocarcinomas and grafted them into adult mice. Following these grafts, he experimentally observed the partial differentiation of the malignant cells into benign cells, which subsequently gave rise to healthy tissues such as muscle. With these studies, Pierce confirmed that teratomas can potentially transform into healthy tissues.
Pierce also highlighted the role of the tissue context in the differentiation processes of embryonic cancer cells: In vitro teratocarcinoma cells can remain stable for up to 28 days, but they begin to differentiate as soon as they are subcutaneously implanted in mice. However, the mechanisms underlying this differentiation remained unclear. Pierce himself explained: ‘The data do not rule out a mesenchymal induction in the embryonal carcinoma as a result of some stimulus originating in the tissue culture environment. (…) We have observed tissue genesis from embryonal carcinoma, but the inductive stimuli for most of these morphogenetic events were not apparent from the study. (…) Differentiation of a loose reticular type of mesenchyme from embryonal carcinoma occurred when embryonal carcinoma was overlaid by visceral yolk sac. Whether this effect depends upon direct contact by embryonal carcinoma cells to those of visceral yolk sac or whether a diffusible substance is involved is not as yet known’ (16). Pierce concluded that: ‘This observation, therefore, suggests the development of methods that would direct the differentiation of embryonal carcinoma cells to benign forms as a logical means of controlling this type of cancer’ (16).
In 1974, Brinster provided further evidence on the possibility of inducing a differentiation of tumor cells and on the role of the cell microenvironment in guiding these processes. In his experiments, Brinster injected teratocarcinoma cells from 129/SvJ black agouti mouse testes into a murine blastocyst. Subsequently, he implanted these blastocysts into female albino mice (the blastocysts also came from albino mice) and followed them until birth. He later observed the development of a new, healthy animal and the consequent disappearance of the malignant cells. In one case, he reported the presence of tufts of dark hair on the back of the newborn albino mouse. This most likely revealed the genetic fingerprint of the mice from which the teratocarcinoma cells had been received. Cancer cells lost their malignant traits and participated in the development of the embryo. Commenting on his results, Brinster stated, ‘the embryo environment can bring under control the autonomous proliferation of the teratocarcinoma cells’ (17).
One year later, Mintz and Illmensee (18) confirmed the results of Brinster and were able to analyze the fate of embryonic carcinoma cells in detail. Their experimental work took place in its entirety over eight years: They initially induced a teratocarcinoma on a 129/SvJ black agouti mouse. They then implanted the embryonic carcinoma cells into a testis of a brown C57-b/b mouse that metastasized to the kidney shortly after. The primary testicular tumor was then extracted, and its embryonic tumor cells were transplanted into the intraperitoneal space of the abdomen of another C57-b/b mouse. This gave rise to neoplastic ascites. A series of successive transplants of embryonic tumor cells into other abdomens of syngeneic mice were carried out for seven years until, in 1975, when embryonic tumor cells were injected into a mouse blastocyst of the C57-b/b strain brown that was then implanted in the uterus of an ‘adoptive mother.’ Mintz and Illmensee (18) injected embryonic tumor cells into a total of 280 blastocysts. These were then implanted in the same number of adoptive mothers' wombs. A total of 97 of these were sacrificed and analyzed between the 8th and the 18th day of gestation. The remaining 183 were allowed to reach the term of pregnancy, giving rise to 48 livng mice.
Both fetal and offspring analysis revealed that all animals were healthy and showed no evidence of tumors of any kind. Even more interesting were the results of the analyses on the composition of hair, the type of circulating red and white blood cells, and the protein composition of urine, as well as characteristics of the kidneys, liver and thymus. The teratocarcinoma cells deriving from black 129/SvJ agouti mice had participated in the normal formation of the organs by a ‘mosaic’ integration with the cells of the brown C57-b/b mouse strain. One mouse was then mated and gave rise to healthy offspring, demonstrating that the sperm cells were also normal. The authors highlighted how: ‘In the present experiments, orderly expression of numerous genes (for example, immunoglobulin, hemoglobin, MUP and agouti genes) has occurred in vivo after they had been ‘silent’ or undetected in the tumors for 8 years, as well as in cultures of teratocarcinoma cells’ (18).
Mintz and Illmensee (18) thus advanced the hypothesis that the mechanisms underlying the neoplastic transformation were to be sought not at the level of genes but of their expression processes: ‘The capacity of embryonal carcinoma cells to form normally functioning adult tissues demonstrates that conversion to neoplasia did not involve structural changes in the genome, but rather a change in gene expression’.
A similar concept had already been proposed in 1954 by Grobstein (19) on the occasion of the 13th Symposium of the Society for Development and Growth: ‘The differentiation of such tissues may depend on inductive interactions between embryonic components’. With regard to the aforementioned study, the double recurrence of the term ‘reversion’ associated with tumors is underscored. Grobstein specifically referred to the malignancy of tumors: ‘Reversibility of malignancy of the core cells’ and ‘the results also furnish an unequivocal example in animals of a non-mutational basis for transformation to malignancy and of reversal to normalcy’. Previous studies in fact, used the term ‘differentiation’.
Although this research sounds promising for new therapeutic cancer strategies, no systematic study to improved exploring any ‘differentiation mechanisms’ of cancer cells has followed. Teratocarcinoma is still considered as a curiosity within the world of oncology. Possibly, for this reason, those results were accepted yet ignored as deviations with respect to the general behavior of other types of cancer cells.
At the same time, the ‘gene-centric’ paradigm was successfully entering cancer studies as the first chemotherapy approaches gave promising results. Cancer reversibility data from teratocarcinoma models need to be verified on other tumor cell lines. This, in order to substantiate the rationale of ‘differentiation treatments’ as a possible cancer treatment approach.
Virus-induced tumors and reversion
Significant evidence of non-teratocarcinoma tumor reversion came from the study of Ian Macpherson (20). He focused on virus-induced tumors, specifically the Rous sarcoma virus (RSV). This virus, which was discovered by Peyton Rous in 1911, can induce sarcoma in the cells that it infects.
Macpherson's series of experiments demonstrated that RSV-infected cells that had become cancerous could undergo ‘reversion’ (using his term) after repeated in vitro transplantations of cell cultures. Under the best experimental conditions, Macpherson was able to obtain a reversion of 19% of the cells after three weeks of culture and 98% after eight weeks of culture. It is important to note that Macpherson used the term ‘reversion’ when he observed that tumor cells resumed their orientation in an orderly manner as in normal tissues, i.e. re-acquired a ‘normal’ phenotype (21).
An important contribution comes from Pollack et al (22) who in 1968 isolated for the first time spontaneously reverted cancer cells. They obtained cancer cells by infecting NIH3T3 cells with SV40 or Polyoma virus. It was observed that some of these cells underwent a spontaneous phenotypic reversion. The cells lost their malignant traits and acquired a flat morphology. Therefore, these cells were named ‘flat revertant’. Subsequently, the ‘flat revertant’ cells were selected by eliminating the non-revertant cells with FUdR (22). This represents a very interesting model to study the mechanisms underlying reversion of cancer cells.
Being inspired by previous studies carried out in the 1940s (23) In the 1980s, Dolberg and Bissell (24) carried out a study on chicken sarcomas, which confirmed the differentiation and protective potential of the cell microenvironment against tumors. Early chicken embryos were infected with RSV. This infection, which gave rise to sarcomas in adult chickens, did not lead to any malignant degeneration between the embryonic cells, even though the virus was active inside them (24). These experiments also highlighted the potential anticancer role of embryo microenvironment.
Tumor reversion in plants
At the turn of the 1950s and 1960s, Armin Braun, a researcher in plant genetics at the Rockefeller Center, developed an experimental method to differentiate plant tumor cells. Specifically, Braun worked on teratomas of tobacco. The experiment aimed to understand how different structures and tissues could emerge from a single tumor cell. Braun highlighted how specific environmental factors involved in cell growth and division such as auxins, and in cell division such as cytokines, were able to determine cellular differentiation (25,26). Interestingly, these mechanisms, which control growth and differentiation, play a role in both germ and cancer cells. In subsequent studies, Braun observed that it was possible to transform a malignant phenotype into a benign one by cultivating plant tumor cells in contexts with no auxins or cytokines, which are the metabolites necessary to support ‘tumor metabolism’ (26).
A series of experiments involving sequential grafts of teratomas on healthy plants further demonstrated the possibility of transforming a malignant tumor phenotype into a benign one. After growing clonal teratomas, Braun grafted them onto the canes of healthy plants. The teratomas proliferated but diminished their degree of malignancy. Braun then took cell samples from these second tumor masses and grafted them onto other healthy plants. He repeated these steps three times until the teratoma disappeared completely. The result was normal plant growth of the plants. Once planted, the seeds gave rise to new, perfectly normal tobacco plants (26).
Through his experiments, Braun demonstrated that cancer cells are endowed with plasticity and that it is possible to grow a healthy plant from a cancer cell: ‘Results of this study indicate that the capacity of teratoma tissue of single cell origin to organize is a reflection of the inherent potentialities of pluripotent tumor cells (…) clones of teratoma tissue of single-cell origin developed organized structures (…) a controlled recovery of crown-gall tumor cells could be accomplished’ (26).
Based on these results, Braun advanced the hypothesis that there may be a hierarchical relationship between mutations at the level of genes and control by the cytoplasmic and tissue context: ‘When tumor shoots derived from tumor buds were forced into rapid growth by a series of graftings to healthy plants, they gradually recovered and became normal in every respect. These results suggested that the cellular alteration in crown gall did not involve a somatic mutation at the nuclear gene level since heritable changes of that type are not generally considered to be lost as a result of rapid growth’ (26).
Since plant dynamics differ from those of animals, it is difficult to translate observations from one area to another. However, the idea that a new fertile plant with ‘healthy’ seeds can be originated from a tumor cell remains stimulating. Anyway, this research highlights two aspects that have already been observed in animals: a) embryonic tumor cells can differentiate, and b) the microenvironment plays a fundamental role in guiding the differentiation processes.
In vivo model of spontaneous cancer regression
In the 1940s, Rose (27) and Wallingford (28) documented cases of in vivo renal tumor regression. They took Lucké kidney tumors from frogs and implanted them on the limbs of some salamanders that were undergoing a regeneration process. Following these grafts, Rose and Wallingford observed the arrest of tumor growth and the subsequent differentiation of cancer cells into cartilage, muscle and connective tissue cells. However, they could not define whether the differentiated cells came from the frog and therefore from the kidney tumor or the salamander. This prevented them from establishing with certainty whether the observed process was a differentiation process of tumor cells or simply an arrest of cancer cell proliferation (27,28). It should also be noted that Lucké renal tumor has a viral origin. Therefore, it could not be a valid model for tumors caused by carcinogenic chemical agents.
In those same years, Gersch (29) compiled a list of the various types of tumors observed on different animal species and their frequency. The list demonstrated that animals with high regenerative capacity have a very low rate of tumor onset. An association between regenerative processes and protection from tumors thus became conceivable (29).
A decade later, Seilern-Aspang and Kratochwil combined the observations of Gersch with the hypotheses advanced by Waddington (30) and Needham (31) according to which tumors might emerge from a loss of control of cell differentiation by the context. More precisely, Waddington and Needham took into consideration the so-called ‘morphogenetic field’, that is, a biological organizational scheme that emerges from the integration of biological signals, for example, cell-cell interactions, and biophysical constraints, for example, forces related to the stiffness of a given tissue or diffusion processes that alter the cellular behavior. Starting from these premises, Seilern-Aspang and Kratochwil (32) designed an experimental model aimed at studying the processes of carcinogenesis and tumor plasticity in newts. These animals feature a highly regenerative power of the limb, so they must host strong morphogenetic processes. The authors worked on newt epithelial tumors induced through an exposure to carcinogens on different sites of an animal's body. In this way, they overcame the experimental limitation of the work of Rose and Wallingford on the viral origin of cancer.
Following the exposure to carcinogens, the animals developed tumors that progressively acquired malignant characteristics. These evolved from an expansive to an infiltrative and a metastatic phase. This behavior confirmed the tumor nature of the processes induced with carcinogens.
By monitoring the spontaneous evolution of tumors, a strong tendency toward spontaneous regressions was observed. The frequency of these regressions varied according to the anatomical areas where the tumor had been induced. In order to verify whether a differentiation of cancer cells had actually occurred with their consequent reintegration within healthy tissues, histological sections were made on both partially regressed and completely regressed tumors. It emerged that the tumor cells had undergone differentiation and integration into normal tissues. In some cases, the differentiated tumor cells had abnormal structures-but not of a cancerous nature. The authors also noted that the spontaneous resolution of metastases occurred almost simultaneously with the resolution of the primary tumor (32). These results made it possible to advance the following hypothesis: The natural processes of tissue regeneration were also able to induce and guide the differentiation of cancer cells.
A further interesting result was reported by McMichael (33) who observed a partial tumor regression on a rabbit skin papilloma following the administration of vitamin A. The role of vitamin A as a potential anticancer agent was further studied by Saffiotti et al (34) in mice exposed to the carcinogen benzopyrene. Vitamin A-administered mice exposed to benzopyrene tended to develop far fewer squamous lung tumors. Tumor regression following vitamin A administration was also observed by Davies on murine skin papilloma (35).
In the early 1990s, Coleman et al (36) studied the fate of two different tumor cell lines resulting from the neoplastic transformation of the liver epithelial cell line WB-F344. The two types of tumor cells (GN6TF and GP7TB) were labeled with the retrovirus BAG2 and the PKH26-GL dye in order to easily identify them in vivo through histochemical techniques. When injected subcutaneously, these two tumor cell lines gave rise to aggressive cancer in 100% of the animals within 18 to 21 days. This confirmed their high malignant potential. When, on the other hand, they were injected into the liver tissue of mice, their aggressiveness was considerably attenuated. The GN6TF line did not produce any type of tumor. Instead, it morphologically differentiated and integrated into the liver tissue. The GP7TB line, on the other hand, gave rise to highly differentiated and not very aggressive intrahepatic tumors.
Following these results, it was hypothesized that the liver microenvironment may exert differentiation action on some types of cancer cells by eliminating or reducing the tumorigenic potential: ‘The apparent complete morphological differentiation of BAG2-GN6TF cells suggests that the microenvironment of the liver not only suppresses the ability of this particular tumor cell line to form tumors but also stimulates them to integrate into hepatic plates and differentiate into hepatocytes. By contrast, BAG2-GP7TB cells form tumors in the liver that are more highly differentiated morphologically than are tumors that form at subcutaneous transplantation sites, suggesting that the regulatory influence of the liver parenchyma can induce partial reversion of the transformed phenotype without complete suppression of tumorigenicity’ (36).
Interestingly, Coleman et al (36) used the term ‘reversion’ (in this case, ‘partial reversion’) to describe the phenotypic transformation of cancer cells. The shared elements of this different experimental evidence obtained on animal models require further investigation. First, the regenerative properties of tissues (even liver cells are characterized by a high regenerative frequency) can exert a control action on cancer cells. This action is very likely to be exerted by the cellular microenvironment as a result of regenerative and morphogenetic processes. These observations are in accordance with the results obtained from the transplantation of tumors within the blastocyst. Generative/regenerative processes such as embryonic development appear to have the ability to regulate the development of cancer cells. It should be noted that these experimental works did not find a return of cancer cells to their original stage. Rather, the loss of their malignant characteristics and the integration within the healthy tissues was observed. Therefore, the term reversion should be intended only as related to the benign-malignant phenotype.
Embryo microenvironment and cancer cell differentiation
As already aforementioned, Barry Pierce was the first to set up a systematic study on the role of the embryonic microenvironment in determining the differentiation of cancer cells. ‘Alternative to cytotoxic therapies are desperately needed for the treatment of carcinoma with metastases. I would propose the direction of differentiation of malignant to benign cells as the most promising alternative’ (37).
Several experimental observations have highlighted how embryonic tumor cells could undergo differentiation if injected into a blastocoele. On the contrary, no regulation took place if the cells were injected into the perivitelline space (38–40). According to these observations, the embryonic microenvironment and, more specifically, the injection site of tumor cells, could have differentiation potential on tumor cells. This has led to the deduction that only some types of the cellular microenvironment are suitable for inducing a differentiation of tumor cells.
Pierce et al (39) took a step further when tried to inject different types of cancer cells such as leukemia, sarcoma and neuroblastoma into the blastocyst. With leukemia and sarcoma cells, no control action was observed on the development of tumor cells. With neuroblastoma cells, only a slight control action was observed (39). These data led to the hypothesis that the control action on cancer cells could only be possible if the embryo had already developed the respective ‘healthy’ phenotype of cancer cells. Since the cells are still undifferentiated at the blastula level, it was decided to inject neuroblastoma cells at a more advanced stage of embryonic development, specifically the neurula stage during which nerve tissues are formed. The study was carried out by Podesta, a collaborator of Pierce. The neuroblastoma cells into a mouse embryo at 8 ½ days of development and observed normal development of the embryo in 80% of cases. These results indicated that the tumor cells had been regulated and directed toward physiological differentiation processes (41,42). These data appear to demonstrate that, depending on the specific embryonic developmental stage, it is possible to regulate different types of cancer cells.
Subsequent studies confirmed the following hypothesis. Leukemic cells injected in the blastocyst did not undergo any differentiation. However, if injected into the placenta of a 10-day murine fetus, they underwent a correct hematopoietic maturation with consequent normal development of the embryo (43). The role of the embryo microenvironment in preventing tumor growth and promoting a phenotypic differentiation of sarcoma in chicken was also highlighted by Dolberg and Bissell (24), as previously described.
Further confirmation of this rationale was provided by Gerschenson et al (44). It was managed to obtain a renormalization of B16 strain melanoma cells following implantation within embryos in a uterus. Both the identification of the correct phase of embryonic development and the correct implantation site were crucial. In consideration of the precise moments in which embryogenesis passes through the phase of formation of melanocytes, the melanoma cells were implanted both in the skin of the back of a mouse fetus at 10 days of development and on the tips of the limbs in formation at the 14th day of embryonic development. These moments correspond to the phases in which melanocytes differentiate at those specific anatomical sites. Consistent with the starting hypotheses, high differentiation rates of melanoma cells were observed with consequent normal development of the embryos. As a control, melanoma cells were implanted on the skin of the back of 14-day-old mice and immediately after being born. In these cases, no significant differentiation rates were recorded, but cancer developed in between 70 and 80% of cases. Gerschenson commented: ‘We have long proposed that cancer is a problem of developmental biology and that an embryonic field capable of differentiating a stem-cell lineage should be able to regulate its closely related kind of cancer. If true, understanding the mechanism of differentiation could lead to non-cytotoxic cures for cancer’ (44).
Within these experiments, an issue remains unclear: What drives cancer reversion processes, the diffusible substances present in the fluids of the cellular microenvironment or the physical contact between cells? A first attempt to investigate this question was made by Pierce et al (45). Embryonic carcinoma cells were exposed exclusively to the fluid extracted from the blastocoele. In this case, no differentiation of the tumor was observed. Rather, it occurred with the graft inside the blastocyst. Thus, it was concluded that it was the cell-cell contact that played the fundamental role in determining tumor differentiation (45). These conclusions were not entirely correct.
In fact, ten years earlier, DeCosse et al (46) had succeeded in obtaining a differentiation of murine mammary adenocarcinoma cells (of the BW 10232 line) by exposing them to mammary embryonic mesenchyme. Commenting on their study, it was hypothesized that: ‘An agent or agents which was inactivated by formalin, probably stable to heat, and capable of traversing a 0.45-µm Millipore filter initiated several morphologic and functional changes in the mammary tumor compatible with differentiation: Namely, development of tubules; interruption of DNA synthesis; changes in nuclear and cytoplasmic morphology; and appearance of a matrix tentatively identified as containing acid mucopolysaccharides’ (46). This way, potential candidates in diffusible substances were identified as inducing causes of tumor differentiation.
A confirmation in favor of the hypothesis on ‘diffusible substances of the microenvironment’ was presented in 1988 by Biava et al (47). The suppression of tumor development on mice lungs primarily induced by homogenates of pregnant murine uteri was observed (47). Subsequent in vitro experiments on different lines of human tumor cells (glioblastoma, melanoma, renal adenocarcinoma, breast cancer, lymphoblastic leukemia), treated with extracts of zebrafish embryos taken during the different stages of cell differentiation and before the gastrulation processes, have achieved a reduction in tumor proliferation on all cell lines. No results were obtained when the embryonic extracts were received in the phases following gastrulation-phases in which proliferative activity prevails over differentiation (48,49).
The zebrafish embryo model was also used by Lee et al (50) in 2005. Human melanoma cells were implanted inside zebrafish embryos in their early stages of development. In this case also, the suppression of the malignant tumor phenotype and the birth of perfectly healthy fish were observed. The following year, Cucina et al (51) treated human colon cancer cells (Caco2) with protein factors extracted from zebrafish embryos in the pre-gastrulation stage. Their experiments confirmed the results that had been previously obtained (49,50) and demonstrated a reduction in tumor proliferation. Cucina et al (51) were able to describe the induction of apoptotic processes mediated by embryonic factors through the activation of mechanisms independent of p53 and linked to the pRb system/E2F1. The synergistic effect of these specific embryonic factors was also demonstrated in vitro when, in the treatment of colon cancer, they were combined with 5-Fluorouracil (5-FU).
In general, the aforementioned study (51) substantiated Pierce's (37) initial hypotheses, i.e. that the embryonic microenvironment has specific characteristics that have yet to be identified and make it able to control the proliferation of cancer cells directing them toward a path of differentiation and normal phenotypic maturation: ‘(…) It is our hypothesis that there must be an embryonic field capable of regulating every carcinoma. Study of how the embryo regulates malignant cells appears promising as an alternative to cytotoxic therapy for carcinoma’ (37).
Since it was functional in structuring this model, Pierce took up the concept of morphogenetic field (30,31). According to Waddington, cancer emerges as a consequence of the loss of control of the morphogenetic fields on cells (30). These concepts were taken by Pierce and Johnson (52,53) and were both applied to the morphogenetic processes of adult tissues and to the morphogenetic processes during embryo development. From this perspective, it can be said that the morphogenetic fields that guide the processes of embryogenesis are also capable of exercising control over tumor cells. It is therefore possible to advance an interpretation of the neoplastic process according to the criteria of developmental biology. In this sense, the tumor can be described as the consequence of the loss of control over the cells by the morphogenetic fields (37) and not just as the result of a progressive accumulation of genetic mutations. Clearly, the question remains open as to whether this ‘escape’ from the constraints of the morphogenetic fields depends on changes in the individual cells or in the microenvironment. It cannot be excluded that the problem should be analyzed from a systemic point of view, i.e., considering the cell-microenvironment equilibrium as an integrated system and therefore hypothesizing that both causes may intervene in the processes of escape from the control of morphogenetic fields.
A very interesting fact that emerges from these different studies is that the various types of cancer cells are selectively sensitive to those specific embryonic microenvironments present in the stages of development during which the corresponding types of healthy cells are differentiated and organized. More simply, neuroblastoma cells are sensitive to the microenvironment taken during the neurula stage, epithelial tumor cells are sensitive to the microenvironments present during the corresponding development stages. This observation made it possible to predict in advance which microenvironments to select to re-normalize specific types of cancer cells.
These concepts are in contrast with the gene-centric explanation of the tumor that were developing and consolidating during those same years, absorbing most of the public and private research funds. As Kenny and Bissel (54) explained, this is most likely why the research on tumor differentiation/reversion, despite convincing and promising results, did not arouse the interest of the international scientific community.
The reversion of acute promyelocytic leukemia (APL)
Despite the lack of interest in differentiation approaches, it was in those years that the first clinical success was achieved: The treatment of APL, a disease that occurs with bleeding and low platelet counts due to the reduced ability of the bone marrow to produce platelets. Today, the current clinical protocol envisages inducing a synergistic effect with a combination of retinoic acid and arsenic, and about 90% of APL patients achieve complete remission (55).
At the basis of APL is a genetic mutation: The gene that codes for the nuclear retinoic acid receptor alpha (RAR-α) and the one that codes for a protein called promyelocytic leukemia protein (PML) blend together. The result of this fusion is the formation of a hybrid protein, PML-RAR-α, which inhibits the functioning of the retinoic acid receptor (56). Following this inhibition, the hematopoietic differentiation processes stop, and poorly differentiated leukemic cells accumulate (57). In parallel, the PML-RAR-α hybrid protein activates genes that maintain the stem cell phenotype and suppress the DNA repair genes. The resulting mutated phenotype promotes tumor progression (58).
Retinoic acid was initially studied as an agent capable of promoting the differentiation of teratocarcinoma cells in vitro (59). However, the results were not as valid in vivo (60). Subsequently, it was observed that retinoic acid could induce a complete differentiation of leukemic cells into granulocytes in vivo (61). Granulocytes were then digested by stromal macrophages, allowing for a rapid elimination of malignant cells and the complete remission in most patients (62).
In fact, treatment with retinoic acid favors the initiation of the degradation processes of the hybrid protein PML-RAR-α (63) and stimulates the differentiation and possible apoptosis of APL cells (64,65). APL remains the only cancer treatment capable of inducing the reversion of the disease and represents a valid ‘proof of principle‘ of the clinical application of reversion.
The aforementioned rationale identifies the essential element in the embryonic microenvironment. Here, instead, a radically different mechanism is at work, that is, the administration of one single substance, trans-retinoic acid. Indeed, Gootwine et al (43) had documented the differentiation of leukemic cells following their grafting into a mouse embryo at the 10th day of development. However, the clinical translation of this approach is very problematic, as it would involve administering a mouse embryo extract to patients.
These results on APL raise some interesting questions that require further investigation. First, are there differences in the differentiation/reversion mechanisms between liquid tumors and solid tumors? Is it possible to hypothesize different mechanisms, not necessarily superimposable, which lead to tumor differentiation/reversion and may involve either the microenvironment or single substances? Both hypotheses, that of the microenvironment and that of individual substances, are worthy of research.
Clinical evidences of tumor reversion
Unlike liquid tumors, such as APL, solid tumors present a greater biological complexity (66). For this reason, the related clinical results are most likely not comparable to those achieved for the treatment of APL (67).
However, numerous clinical studies over the decades have documented cases of the spontaneous regression of tumors. Apparently, these processes are comparable to those observed on salamanders by Seilern-Aspang and Kratochwil (32). The first clinical documentation of spontaneous cancer regression dates back to the 19th century. In 1918, Rohdenburg published an analytical review of 302 cases of spontaneous regressions, with 70 of them as certainly valid (68). Cases of spontaneous cancer regression were reported periodically in medical literature, for example, one related to neuroblastoma (69) or lung metastases deriving from renal cancer (70).
The first monograph on spontaneous cancer regression appeared in 1966. Everson and Cole (71) presented 176 well-documented cases of spontaneous cancer regressions that had been published between 1900 and 1964. In their monograph, criteria for ascertaining the diagnosis of both the disease and its regression were also proposed, which involved histological and radiological documentation. To be valid, the regressions must have occurred without specific therapies, except those whose ineffectiveness was clear (71). This analytical review highlighted that most of the regressions were recorded on four main types of tumors, namely renal tumor, choriocarcinoma, neuroblastoma and malignant melanoma.
In light of this research, the American National Cancer Institute sponsored a conference on the topic of spontaneous tumor regression in 1974. A monograph was later drafted (72). Further significant research on the subject was conducted by Challis and Stam in 1990 (73). All spontaneous regressions that had been reported between 1900 and 1987 were analyzed, detailing the progressive increase in regressions of lymphomas and kidney tumors (73). In 1993, O'Regan and Hirshberg published a bibliography of all the reported spontaneous regressions of both malignant and benign tumors (74).
In 1998, Papac (75) published an investigative work about the possible mechanisms underlying spontaneous regressions. It was clarified that the spontaneous regression of cancer means a complete or partial disappearance of the malignant tumor in the absence of therapies capable of inducing anti-neoplastic effects. It was also pointed out how numerous patients had experienced relapses after a first regression, which meant that spontaneous regressions were not always stable and could not be always associated with recovery.
The main mechanisms proposed by Papac involved the immune system, hormonal changes, the necrosis of tumor cells, trauma and changes in the vascular system. Mechanisms related to apoptosis and cell differentiation were also proposed. Tumors were classified based on the reported frequency of regressions and the four most recurrent were highlighted, which are kidney cancer, neuroblastoma, breast cancer and melanoma. It was thus partly confirmed what had emerged from the studies of Everson and Cole. In addition, liquid tumors, such as leukemias and lymphomas, were reported.
Another observation that emerged from Papac's study is that the rarest tumors were those in which spontaneous regressions have occurred more easily. On the contrary, spontaneous regressions have occurred more rarely in the most frequent tumors. Not only the type of tumor but also the site of its onset was related to the frequency of regressions. For example, spontaneous regression occurred more easily in the lungs or on the skin. This suggested that, perhaps, certain microenvironment factors facilitated the regression and, more generally, that there were mechanisms for the endogenous regulation of tumor growth processes (75).
Further data came from a series of molecular biology studies on neuroblastoma cells that had experienced reversion: In these cases, a decrease in telomerase activity was observed. Similar studies on retinoblastoma cells instead highlighted how regressing masses also demonstrated an increase in DNA hypomethylation. Neuroblastomas, together with testicular germ cell tumors and acute leukemias, were those in which regression due to cell differentiation was most frequently recorded (75).
Finally, the regression of metastases has been reported after the surgical removal of primary tumors. This fact suggested a sort of paracrine mechanism by virtue of which the primary tumor promoted the growth and proliferation of metastases at a distance (75). These cases of spontaneous regression confirmed the hypothesis according to which the tumor is not a completely irreversible disease. However, identifying a strategy for developing differentiation treatments have remained difficult. The only structured clinical trials with a clear protocol based on differentiation therapy were those related to APL.
A prospective clinical trial on liver cancer published in 2005 is therefore remarkable. This study reported some cases of tumor reversion following non-chemotherapy treatments. This randomized clinical study involved 179 patients with advanced hepatocellular carcinoma and no chance to undergo any type of therapy. One of the two groups of patients was administered, in the form of compassionate care, a zebrafish egg extract. The second group was given no treatments other than conventional pain management for dying patients. The results revealed clear clinical benefits in favor of the group treated with zebrafish egg extract. The study was therefore interrupted in its randomization and, for ethical reasons, the extract was then administered to all 179 patients. The monitoring continued for three years during which 19.8% of the patients had tumor regressions (2.5% were total regressions), and 16% stabilized the progression of the disease. An increase in performance was observed in 82% of patients (76). Clearly these are only preliminary data that need further confirmation.
Molecular mechanisms underlying tumor reversion
Understanding the mechanisms underlying tumor differentiation/reversion is key in developing differentiative treatments. In 1968, for the first time, Robert Pollack isolated spontaneously reverted cancer cells (Pollack, 1968); afterwards Telerman et al (77) further developed this experimental model and produced the first systematic study on the molecular mechanisms underlying tumor suppression by focusing on tumor suppressor genes. In 2002, Telerman introduced the term ‘tumor reversion’. It was defined as follows: ‘The process by which some cancer cells lose their malignant phenotype-and from a molecular point of view-tumor reversion can be defined at the molecular level, not just as the reversal of malignant transformation, but as a biological process in its own right involving a cellular reprogramming mechanism, overriding genetic changes in cancer, by triggering an alternative pathway leading to suppression of tumorigenicity’ (78).
This concept is very similar to that of ‘reversion of malignancy,’ introduced in 1975 by Mintz and Illmensee (18), who had studied the malignancy of specific tumor phenotypic characteristics. To obtain ‘revertant’ cells, Telerman et al used a selection technique based on the infection of various tumor cell lines with parvovirus H-1. This particular type of virus has the characteristic of selectively eliminating only malignant tumor cells. In this way, virus-resistant tumor cells are selected and have lost their malignant potential. To confirm that they were ‘revertant’ cells, these cells were subsequently implanted in mice to observe the stable reduction of tumorigenicity.
The comparison of the different genetic profiles of various leukemic and breast cancer cell lines let a common group of genes stand out among all of the different cells that had undergone a phenotypic reversion of malignancy. Depending on the type of tumor, this group of genes had different expression ratios. However, the ‘overall variable’ of their genetic network seemed constant among the different types of tumors.
Of the nearly 300 genes involved in the reversion process, Telerman was able to identify the main ones, namely, seven in absentia gene (SIAH1), presenilin 1 (PS1), tumor suppressor-activated pathway (6TSAP6) and translationally controlled tumor protein (TCTP). Two of these genes, SIAH1 and TSAP6, are upregulated in the reverting tumor cells. The other two, PS1 and TCTP, are instead repressed (78–80).
More specifically, SIAH1 is a target gene of the p53 protein. This protein plays a crucial role in apoptosis processes. SIAH1 promotes the degradation of β-catenin, which is involved in cell adhesion processes and mitigates the activation of the signal cascade mediated by wingless-related integration site (Wnt). When activated, these signals contribute to stimulating cell proliferation. Overactivation of this gene, therefore, favors tumor suppressor mechanisms. The overexpression of the TSAP6 gene also favors the activation of mechanisms that promote apoptosis. Conversely, when downregulated, the PS1 gene leads to tumor reversion. This is because PS1 in itself exerts an anti-apoptotic action; therefore, its downregulation silences this inhibitory action. The TCTP gene also plays an important role in reversion when downregulated. Of all genes, this is perhaps the most involved in the stabilization and promotion of tumor growth processes, therefore a reduction in its expression levels favors tumor reversion (79).
TCTP was significantly downregulated in all ‘revertant’ cells compared with malignant tumor cells. This protein is widespread among most eukaryotes, and its functioning is associated with tumor growth and acute allergic responses (81). At a physiological level, TCTP has a pro-tumor action because it promotes the growth and stabilization of the cytoskeleton, thus favoring the spread and invasiveness of tumor cells. It also blocks apoptosis by inhibiting p53.
To verify whether this protein had actually played a central role in promoting tumor reversion processes, the researchers inhibited its expression through antisense cDNA. In fact, they could then observe the suppression of the malignant phenotype. TCTP could therefore be a target for possible reversion inducing drugs. However, the process is complex and, as Telerman explains: ‘The gene expression profile suggests that it is not the processes per se of cell cycle arrest, apoptosis and terminal differentiation, that matter here, and that provide by themselves the framework for reversion. It is rather a ‘reorganizing’ function of all these processes as a form of rerouting and trigger of the whole machinery that enables the tumor cells to quit the malignant pathway, even bypassing mutant or wild-type p53’ (78).
Consistent with this hypothesis is the research by Proietti et al (82) who, in 2019, managed to induce phenotypic reversion on two different breast cancer cell lines (MCF-7 and MDA-MB-231) by using a zebrafish embryo microenvironment. They obtained the phenotypic reversion of cancer cells and highlighted some mechanisms through which the reversion is activated. Specifically, these were a remodeling of the cytoskeleton and a downregulation of TCTP. Apoptosis and phenotypic differentiation immediately followed (82). Subsequent studies confirmed the same results with different cancer cell line cultures (liver cancer, colon cancer and glioblastoma) and different embryo models, specifically trout embryo (Bizzarri, forthcoming).
An interesting work by Weaver et al (83) highlighted the role of integrins and cell adhesion processes in triggering phenotypic reversion. They used human breast cancer cells cultured in 3D with inhibitory β1-integrin antibodies. The reversion of tumor phenotype occurred following the inhibition of cancer cells β1-integrins and the subsequent re-normalization of adherens junction assembly. This showed that, despite genetic mutations remaining in cells, it is possible to re-establish normal phenotype and normal tissue morphology. Moreover, from the aforementioned study emerged the role of the basement membrane in controlling cell proliferation thus highlighting the fundamental role of tissue architecture in modulating the phenotypic expression of cells.
Another investigation focused on the molecular mechanisms of the reversion/phenotypic differentiation of cancer cells was carried out by Hendrix et al (84). This experimental model consisted of exposing two different tumor cell lines (melanoma deriving from the neural crest and breast carcinoma of epithelial origin) to factors secreted by human embryonic stem cells. The result was a loss of malignancy and a consequent reprogramming toward a benign phenotype (84).
Previous research has identified that aggressive melanoma and breast cancer cells exhibit high levels of expression of the gene encoding the Nodal protein, an element that plays a fundamental role in the process of embryonic morphogenesis, thus evidencing possible correlation between embryogenesis and cancer. Furthermore, melanoma and breast cancer also lack the Lefty protein, a natural Nodal inhibitor. This is a molecular signal belonging to the superfamily of transforming growth factor-β (TGF-β). It is expressed in embryonic stem cells and antagonizes Nodal, thus ensuring a balance between the different signals underlying the morphogenetic processes (85).
The concomitant overexpression of Nodal and the absence of its inhibitor Lefty, therefore, gave rise to an abnormal and unregulated behavior in the tumor cells. As a result, the cells underwent uncontrolled proliferation.
Consistent with this hypothesis, it was observed that exposing these two tumor cell lines to the microenvironment of Lefty-rich embryonic stem cells could induce an inhibition of Nodal and a consequent reprogramming of the tumor cells. These thus reacquired a benign phenotype. Cancer cell lines were also exposed to microenvironment soluble factors from other types of stem cells, such as those deriving from amniotic fluid, umbilical cords and bone marrow. All these types of stem cells do not produce Lefty and, consistent with the working hypothesis, their microenvironment was not able to induce a reprogramming of tumor cells (86). This mechanism, similar to the one identified by Telerman on TCTP, represents a possible target for new therapeutic strategies on cancer.
In general, this research confirmed the presence of profound similarities between the processes of embryogenesis and those of tumor reversion and differentiation. In particular, the authors explain how: ‘The phenotype of stem cells and cancer cells is profoundly influenced by the microenvironment. During embryogenesis, precursor cells are specified to particular fates through the delivery of signaling molecules, and malignant cells similarly release and receive cues that promote tumor growth and metastasis’ (86). Among these signals, the main ones are those belonging to the superfamilies of Notch, Wnt, TGF-β and Nodal (87).
It must be stressed that all of these are epigenetic processes. They control the gene expression through environmental signals. These signals interact with DNA and its expression, for example, via methylation and acetylation. Indeed, DNA methylation and histone modification are among the most known epigenetic events. When tumor promoters and tumor suppressors involve genes, these events can, regardless of the presence of mutations, lead the behavior of the cell toward a tumor path. This is why interest in micro-RNAs (miRNAs) is growing. Micro-RNA is a type of non-coding RNA that can play an important role in guiding the epigenetic modulations of DNA (88). A single miRNA can interact with hundreds of thousands of target proteins and promote systemic action (89).
To investigate this hypothesis, an experimental model was developed in which C8161 line melanoma cells were exposed to the microenvironment of human embryonic stem cells. The expression profiles of miRNAs within the tumor cells were studied before and after exposure to the embryonic microenvironment. MiRNAs were thus identified, some of which were upregulated in melanoma cells, specifically miR-302a. Others, on the contrary, such as miR-27b, were downregulated (90). These miRNAs were also associated with embryogenesis processes, thus confirming the thesis according to tumor reversion processes have strong similarities with embryo genetic processes (91).
The aforementioned study also highlighted a connection between miRNAs and Nodal regulation. In fact, it identified a second Nodal inhibition mechanism that exploits the Nodal-Notch4 axis. This axis is key in embryo genetic processes: It drives the formation of the right-left axes (92). Lefty promotes an increase in miR302a levels which, in turn, silences the Nodal and Notch4 signaling circuit. The result is a loss of the malignant phenotype by melanoma cells (90).
Cell cycle control is key in maintaining a correct balance between cell proliferation and differentiation. Precisely inside the cells, there are proteins called cyclin-dependent kinases that have the task of blocking uncontrolled cell proliferation. For this reason, they are also called ‘gatekeepers‘ since, in correspondence with the different checkpoints of the cell cycle, they block abnormal cells (93). Clearly, cancer cells escape these controls and continue to proliferate, completing the entire cycle undisturbed. A possible therapeutic strategy for cancer, therefore, could aim at restoring the correct functioning of the various cell cycle checkpoints or forcibly blocking the tumor cells in the initial G1 phase of the cell cycle. In 2009, Giuffrida et al (94) developed an experimental model exposing different cell lines of human epithelial tumors, such as ovarian, prostate, and breast cancer, to substances secreted by embryonic stem cells. The aforementioned study confirmed the ‘antitumor’ activity of the embryonic microenvironment and investigated the cell cycle modification during phenotypic reversion processes.
The aforementioned study confirmed a selective antitumor action of the embryonic stem cell extracts. It also confirmed the selectivity of cancer cells by exposing human fibroblasts to the same factors. In this case, no inhibition was observed. Furthermore, following exposure to the embryonic microenvironment, an important number of cancer cells remained blocked in the G1 phase. At the same time, smaller quantities of cancer cells were detected in the most advanced stages of the cycle, S and G2/M, indicating that the soluble factors produced by embryonic stem cells might have exerted an inhibitory action on the cell cycle, selective toward cancer cells.
In their research, Giuffrida et al (94) characterized these factors as low-weight and thermostable molecules that allow embryonic stem cells to exert paracrine and autocrine actions. These substances could therefore represent a valid therapeutic strategy: ‘Instead of using stem cells themselves for therapy, it will likely be possible to identify and synthesize the specific tumor-suppressing factors secreted by embryonic stem cells, thereby bypassing the practical and ethical issues currently associated with embryonic stem cell therapy’ (94).
Previous literature has already shown that epigenetic alterations are pivotal in the early stages of breast cancer development (95,96). For this reason, it was conceivable that one of the mechanisms through which embryonic extracts could exert their ‘tumor reversion’ action might be the modification of chromatin. In fact, hypermethylation is one of the main silencing processes of tumor suppressor genes. Therefore, the hypothesis to be tested was whether the embryonic extracts were able to demethylate these genes. For this purpose, a series of genes that could be the targets of these actions, such as RARB, CST6, CCND2, GAS2 and CDKN2A, were identified.
Further elements on the possible mechanisms underlying the processes of tumor reversion and differentiation were provided by Allegrucci et al (97). Their experimental model exposed a breast cancer cell line, MCF7, to three different types of ‘embryonic extracts’ from three different sources, namely a Mexican salamander embryonic (Axolotl), frog embryo (Xenopus laevis), and mouse embryonic stem cell. The goal was to both verify the differentiation potential of these extracts on cancer cells and investigate possible epigenetic mechanisms underlying these processes.
The results confirmed the initial hypotheses that both salamander and frog embryo extracts favor the re-expression of these genes, although not always restoring it to normal level. Furthermore, only the extracts received during the initial stages of embryogenesis exerted this action. Extracts received at a later stage did not exhibit this activity. The extracts of mouse embryonic cells, on the other hand, did not give satisfactory results, inducing only the re-expression of the GAS2 gene. However, since reprogramming actions were observed at the cellular level, perhaps the mechanisms worked through other pathways in this case. In general, the most efficient extracts were those from salamanders.
To verify the stability of these epigenetic modifications, the reprogrammed tumor cells were implanted in immunosuppressed mice to be compared with ‘non-reprogrammed’ tumor cells. Tumors from the ‘reprogrammed/reverted’ cells were markedly smaller than the control group eight weeks after transplantation. Even at the histological level, tumors from the reprogrammed cells were more circumscribed, although not encapsulated, and had a lower rate of mitotic division, which indicates a lower degree of malignancy.
These results confirmed that certain embryonic extracts were able to exert a stable reprogramming action on cancer cells. This approach has opened up promising new avenues for research: ‘It will now be important to identify the oocyte-specific molecules involved in this process, and the molecular pathways responsible for the arrest of tumor growth. In our view, the identification of these molecules will provide a rich source of information for the design of synthetic molecules that can be used for pharmaceutical interventions’ (97). Further experiments performed by Saad et al (98) on MCF-7 cells treated with axolotl oocyte extract highlighted more mechanisms involved in reprogramming/reversion processes such as cell cycle arrest associated with upregulated expression of P27 and reduction of RB phosphorylation. An interesting result was the induction of tumor cell dormancy (98).
Proietti et al (82) also obtained the phenotypic reversion of malignant cancer cells. He exposed an aggressive breast cancer line, MDA-MB-231, to zebrafish and trout embryo extracts. His research confirmed the reduction of cell proliferation and highlighted some mechanisms through which the reversion was activated: A remodeling of the cytoskeleton and a downregulation of the TCTP protein that had already been studied by Telerman. This downregulation promoted apoptotic processes and cell cycle stabilization (82). A recent study identified some of the most relevant genes involved in tumor reversion processes, including STAT3, ROCK, BRCA1, SETDB1, MMP9, YBX1, FASN, RARα, RB1, PRKD1, PP14, FAK, and P53 GM2/GM3. In some cases, post-translational mechanisms have been highlighted. These include phosphorylation, methylation, deglycosylation and silyation (99).
All of this experimental evidence provided for a new therapeutic rationale based on epigenetic regulation processes. These processes are increasingly explored in the context of cellular reprogramming. According to this model, the embryo microenvironment can induce the differentiation of tumor cells, consequently promoting a reversion of the malignant phenotype through epigenetic regulation pathways. These are, in numerous cases, similar to those involved in embryogenesis processes. Therefore, the control of tumor development appears to take place at the epigenetic and non-genetic level.
The contribution of cellular reprogramming research in the context of tumor reversion
A big boost to tumor reversion research was provided by the studies on cell reprogramming by Shinya Yamanaka and John Gurdon, granting them the Nobel Prize for medicine in 2012. Takahashi et al (100) demonstrated the possibility of reprogramming totally differentiated somatic cells by transforming them into pluripotent stem cells (iPCS) due to the forced expression of four transcription factors: Oct3/4, Sox2, Klf4 and c-Myc. Following the aforementioned results, the concept of phenotypic reversibility and cellular plasticity became substantiated (101).
Subsequent studies have shown that cellular reprogramming is not strictly necessary to activate the expression of Oct3/4, Sox2, Klf4 and c-Myc. For example, the overexpression of E-cadherin is sufficient to activate the reprogramming with no need for the activation of Oct4 (102). Among these, the main molecular programs involved in cellular reprogramming are the modulation of the Wnt/β-catenin and PI3K/Akt pathway, the downregulation of Prenselina-1 (PSEN-1), Notch and SNAI1, and the increase in the synthesis of E-cadherin (103,104).
Even the biophysical elements of the cell culture environment, such as plate material (glass or graphene) can trigger the reprogramming of fibroblasts in induced pluripotent cells (105). This highlights the importance of the microenvironment and the tissue in determining the processes of cellular differentiation or de-differentiation. In order to grasp the key factors involved in cellular reprogramming, it is necessary to go beyond the study of molecular pathways and look for the tissue level (106).
Despite progress in understanding these mechanisms, clinical application remains far away. It is important to note that when talking about cellular reprogramming, unlike the concept of tumor reversion, the transformation of epithelial cells into mesenchymal stem cells is generally meant. This process is known as epithelial-mesenchymal transition (EMT) and it characterizes neoplastic transformation. These cellular reprogramming processes imply significant changes in cell morphology, such as the acquisition of a rounder shape and the loss of cell-cell contacts typical of somatic cells. All of these morphological transformations favor the acquisition of properties such as motility and invasiveness that are typical of tumor processes (107). This is one of the main reasons why cell reprogramming techniques have not found clinical applications. In fact, the activation of regulatory factors that initiate cell reprogramming often induces tumors (108,109).
Indeed, these data reinforce the hypothesis that cancer may be an error in the process of cell development and differentiation (12) and that, as demonstrated in the past, molecular signals involved in cell differentiation and embryonic development may also exert a role on tumor cells (110,111). Based on these common biological pathways between cellular reprogramming and oncogenesis, various studies aimed at finding effective strategies to apply cellular reprogramming techniques to cancer cells.
Unlike the tumor reversion approach, one of the first strategies developed to induce tumor reprogramming involved the de-differentiation of tumor cells. The goal was their transformation into pluripotent cells that could subsequently re-differentiate into non-malignant somatic cells. Promising results have been obtained on skin tumor cell lines treated with a specific microRNA (miRNA3025) (112) and on murine melanoma cells (113). However, this research is difficult to translate into clinic procedures because the non-selective cell de-differentiation step could favor the onset of tumors in other cells of the body.
In 2013, Rapino et al (114) reprogrammed human lymphoma and leukemia B cells. They transformed them into macrophage-like cells by introducing a transcription factor C/EBPα. Huang et al (115) provided further confirmations in 2014. It was demonstrated how a combination of transcription factors can play an important role in reprogramming human fibroblasts into pseudo-hepatocytes (115). McClellan et al (116) applied the same concept and managed to reprogram lymphoblastic leukemia cell lines using small molecules.
Exosomes also appear to be involved in tumor reprogramming. In 2017, Zhou et al (117) inhibited tumor proliferation in vitro and slowed down oncogenesis in vivo by using exosomes extracted from embryonic stem cells. A further contribution to this innovative cancer treatment approach was obtained in 2018 by Ishay-Rosen et al (118), who managed to transform breast cancer cells into adipocytes by means of a mix of molecules including insulin, dexamethasone and BMP2. Cancer cells were first treated with TGF-β to bring them back to a pluripotent stage. They were then differentiated by exposure to a mix of molecules that directed these cells toward the acquisition of normal phenotype (118).
In 2019, Cheng et al (119) managed to induce direct reprogramming, that is, without going through the reactivation of the pluripotent stage. In addition, in this case, hepatocellular carcinoma cells were exposed to small molecules (a mix of transcription factors). This treatment induced their differentiation into normal liver cells (119).
A recent study on ‘oncogene addicted’ cancer cells provided for further evidence of the possibility of the phenotypic reversion of cancer cells. Li et al (120) obtained tumor reversion by inhibiting specific oncogenes of tumor hepatocytes in xmrk transgenic zebrafish model. By genetic recombination, tumor cells were marked and it was confirmed that tumor hepatocytes morphologically and molecularly converted into normal hepatocytes (120).
Theoretical implications of ‘tumor reversion’
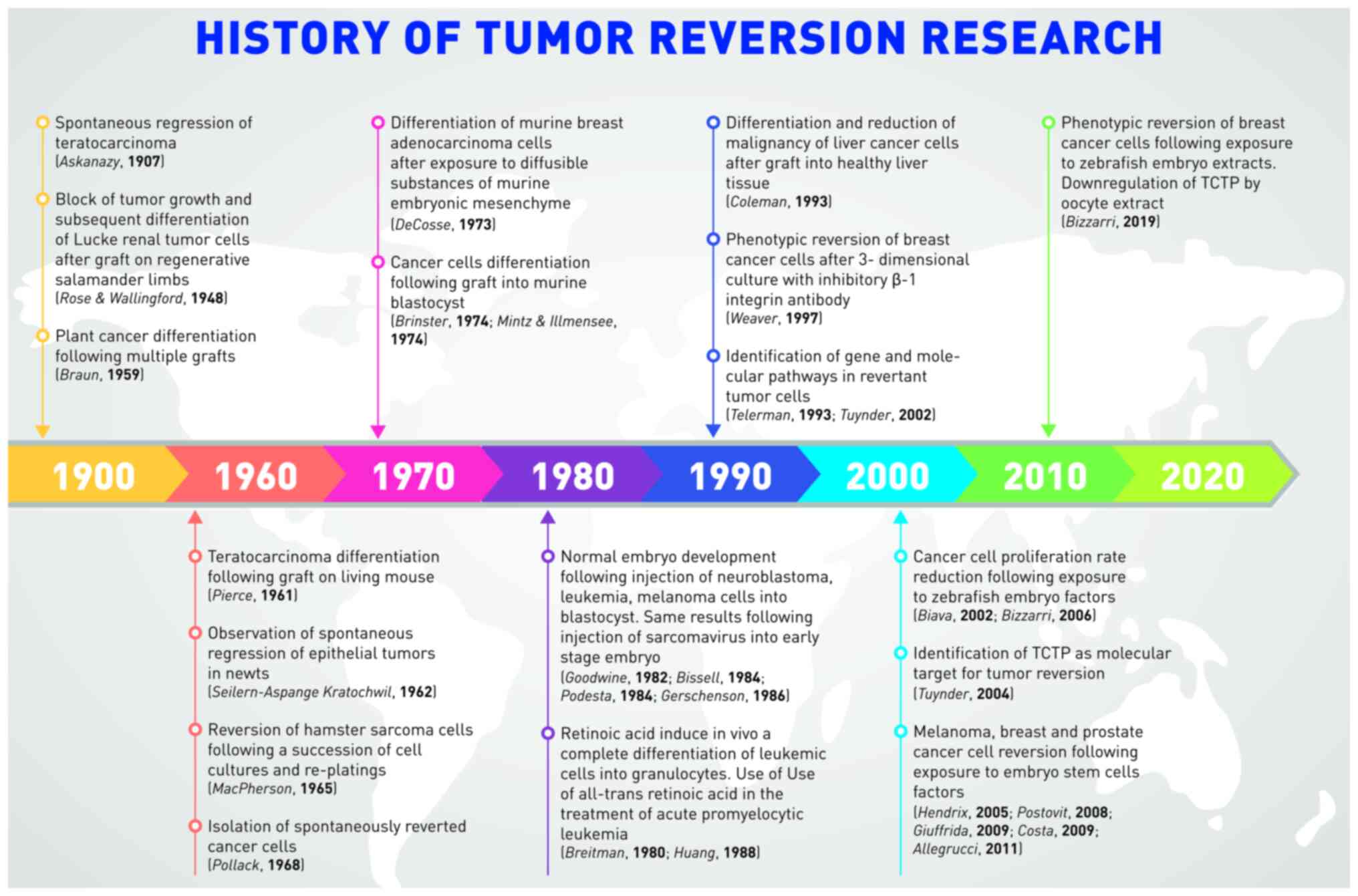
A non-trivial issue deals with the lexicon and the conceptual tools used to describe experimental and clinical observations (121). Four main terms occur most frequently in the literature, referring to ‘tumor reversion’ processes, namely, regression, differentiation, reversion and reprogramming.
The term ‘regression’ appeared in the first half of the 20th century and was mostly used until the 1970s. It mainly refers to the clinical observation of a ‘regressing’ tumor that decreases in mass. In this sense, this term says nothing about the malignant or benign nature of cancer cells. The term ‘differentiation’ was introduced by Pierce and has been used-albeit less frequently-until today. It mainly refers to the characteristics of the cancer cells that differentiate and return to integrate into the tissues, losing their malignancy. Pierce's use of this term took shape from observations on teratocarcinomas in which undifferentiated embryonic tumor cells progressively gave rise to differentiated cells following a kind of embryogenesis processes. This term recalls the conceptual framework of embryonic development and indicates the processes affecting individual cells.
The term ‘reversion’ evokes a markedly more radical concept. It was used for the first time in relation to cancer by Macpherson in 1965, then later structured by Telerman. Macpherson, like Braun, observed the renormalization of cells in their morphology and function and used the term reversion to indicate this recovery of normal characteristics. At the etymological level, the term ‘reversion,’ from the Latin reversio, means to go back and implicitly alludes to a restoration of the original conditions. Such a process in its entirety has not been confirmed experimentally in the sense that the tumor cells have not been induced to ‘go back’ to their original characteristics: in the transformation of the cell from state A (normal) to B (tumor) to C (revertant), it cannot be said that A is equal to C. As aforementioned, Telerman explains this point and the term ‘tumor reversion,’ defined as: ‘The process by which some cancer cells lose their malignant phenotype (…) tumor reversion can be defined at the molecular level, not just as the reversal of malignant transformation, but as a biological process in its own right involving a cellular reprogramming mechanism, overriding genetic changes in cancer, by triggering an alternative pathway leading to suppression of tumorigenicity’ (78). This is very important because it clarifies that the term reversion refers exclusively to the malignant phenotypic characteristics of cancer cells (such as invasiveness and migration) and that the pathways through which cancer cells undergo this transformation can be different. It is therefore more precise to use the expression ‘phenotypic reversion’.
The last term used, which Telerman himself uses in clarifying the definition of tumor reversion, is that of reprogramming; a term that took shape from Yamanaka's work on cellular reprogramming and that is borrowed from the world of information technology. This word refers to the control, programming and activity of the cell's genes and implicitly alludes to an external action, typically human, of the ‘manipulation’ of biological processes. In this sense, it is more difficult to associate the word ‘spontaneous’ with the one of ‘reprogramming.’ It should also be noted that the translation of the concept of programming borrowed from the world of information technology for biology has some application limits (122). Therefore, it should be specified that by cellular reprogramming, it is meant the induction of a controlled transformation of the cell phenotype typically by the path a) de-differentiation, b) re-differentiation, namely in cancer a) EMT-b) MET.
This relative variety in terms reflects the variety of experimental models within which the regression/differentiation/reversion/reprogramming processes have been observed. The various experimental models and results are outlined in Table I.
It is best to find the correct lexical expression for the observed reversion processes so as not to induce, through imprecise terms, erroneous interpretations of experimental data. Nowadays, the literature is still not homogeneous in naming these processes. For the present review, the term ‘phenotypic reversion’ was proposed because it is more precise. It is underscored that some authors use the expression ‘tumor dormancy’ (83,120). This highlights the need for improved comprehension on the nature of these processes as well as their disambiguation. For example, is ‘phenotypic reversion’ a unique process, or do different processes drive cells to different types of benignant states?
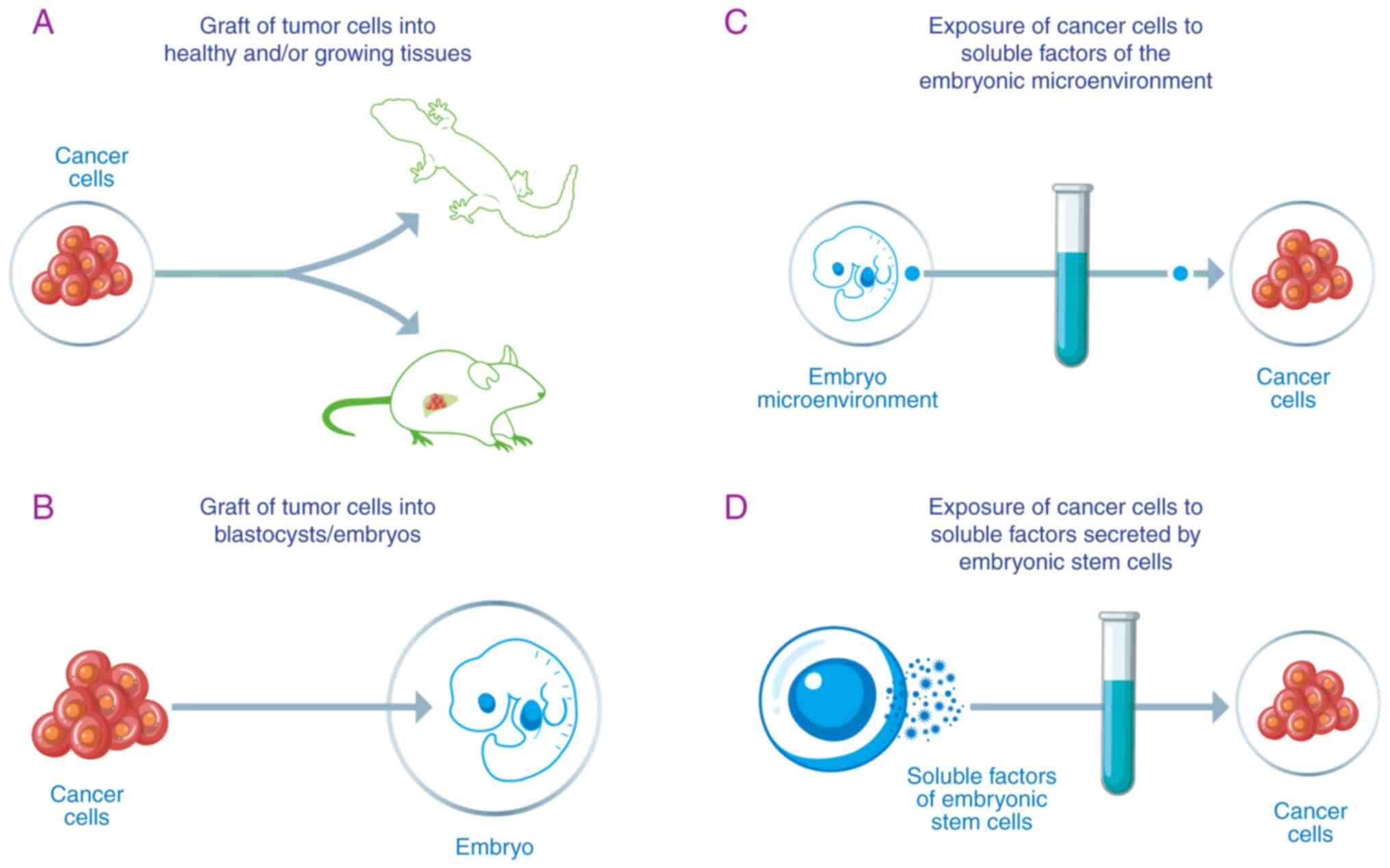
From a deeper analysis of this research, it is possible to identify and classify the following experimental models: i) Transplantation of tumor cells into healthy and/or growing tissues; ii) transplantation of tumor cells into blastocysts/embryos; iii) induction of tumors in healthy animal tissues and monitoring of their evolution; iv) exposure of cancer cells to substances of the embryonic microenvironment; and v) exposure of cancer cells to substances secreted by embryonic stem cells (Fig. 2). Within these models, the various variables taken into consideration are: i) Type of tumor; ii) specific phase of embryonic development in which the tumor is implanted or from which the contained substances are extracted; and iii) anatomical site of tumor implantation.
Conclusions
The present review aimed to present the complex ‘state of the art‘ of this rather unknown field in cancer research. Despite the experimental evidence on phenotypic tumor reversion is based on different experimental models and laboratories, a systematic study program in this field is still missing. Consequently, research programs on cancer research still miss the opportunity to investigate the effectiveness of tumor reversion on cancer therapies. The experimental and interpretative framework is complex and needs a systemic approach to be modeled. The present review only presented a general picture of the ‘state of the art’ and does not try to propose a theoretical model of tumor reversion. However, some conclusions can be advanced on the recurrent elements. First, the fate of cancer cells is not irreversible. This statement implies the ‘proof of principle’ of new therapeutic strategies for cancer. However, it should be noted that ‘non-irreversibility’ does not automatically correspond to reversibility understood as a return to the original state of the cells. It would therefore be more correct to state that the fate of cancer cells is not unchangeable and that cancer cells display relevant plasticity. Second, it is possible to inhibit the phenotypic expression of the malignant characteristics of cancer cells mostly through epigenetic processes, although other mechanisms are likely to participate. This statement implicitly contains the concept of ‘chronicization’ of the tumor. When talking about ‘phenotypic expression,‘ it is necessary to clarify what is meant by phenotype. Specifically, a modification in the behavior of cells that pass from presenting aggressive functions such as the ability to migrate and invade new tissues to have more controlled, and therefore benign, characteristics, is indicated. Third, specific embryonic and tissue morphogenetic fields are capable of exerting a direct action on the phenotypic expression of cancer cells. Within the embryo (at certain stages of development) and in specific healthy tissues, there are substances/mechanisms capable of directing malignant tumor cells toward the re-expression of benign characteristics. This statement implies a reconsideration of the hierarchical relationships between genes and the context of the cellular microenvironment, between genetics and epigenetics. The concept of morphogenetic field is also used to indicate the complex network of biological and biophysical signals that can exert regulatory (and binding) actions on cells. Clearly, in order to describe these processes, new conceptual tools are needed to grasp the complexity and dynamism of a biological network. Fourth, the pivotal role of genes in explanatory models of cancer should be reconsidered and integrated with new conceptual tools such as morphogenetic field, biological network, cell reprogramming and phenotypic reversion. These terms are ‘foreign’ to the conceptual arsenal developed in the reductionist current paradigm within which cancer has been interpreted.
Fifth, new techniques such as single-cell transcriptomics or cell state transition assessment and regulation (cSTAR) may offer important cues on understanding the malignant-benign dynamic processes of cell state transition (123). Sixth, numerous questions remain open, such as those related to the nature of these ‘transformed’ cancer cells, their stability, the biological pathways that lead to these transformations, and the specificity of the interactions between cancer cells and the microenvironment. In fact, not all types of cancer cells respond to embryonic signals and, in light of the heterogeneity of the tumor masses, it is logical to ask how a renormalization of the entire tumor complexity can occur. Furthermore, it is worth mentioning that the most relevant results in tumor reversion experiments derive from the exposure of cancer cells to a raw extract of the embryo microenvironment. In this form, it is difficult to standardize and industrialize the extracts. Issues such as pharmacodynamics, bioavailability and stability are needed for a clinical translation of these approaches.
Seventh, differentiation therapies represent a promising research field for cancer treatments but the way remains long. Several issues should be addressed, not only of a scientific nature, but also technological, industrial and probably regulatory. Do these extracts work in their entirety or there are few active molecules? How should an oocyte extract be juridically classified? What kind of patent is allowed regarding a natural extract? How to measure pharmacodynamics, bioavailability and stability? Eighth, occasionally scientific research opens new ways that are still not integrated under the socio-economic framework and this can represent a further obstacle to the research development. Scientists (and pioneers) should therefore acquire a systemic view not only for their specific research model but also for a wider and more effective integration of their work into society.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
AP carried out the research and classification of the material, wrote the initial manuscript and draw the figures. M Be and MBi contributed to the organization of the work and revising the manuscript. All authors contributed to the manuscript through revisions of earlier versions, and read and approved the final manuscript. Data authentication is not applicable.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
|
Recamier JCA: Recherches sur le Traitement du Cancer, etc. Chez Gabo; Paris: 1829 | |
|
Oberling CH: The Riddle of Cancer. Yale University Press; New Haven: pp. p1961944 | |
|
Oberling C: The riddle of cancer. Yale University Press; New Haven: pp. 26–27. 1946 | |
|
Müller J: Über den feinern Bau und die Formen der krankhaften Geschwülste. G. Reimer, Berlin. Nat Cancer Inst Mnogr Spontaneous Regression Cancer. 1976:441838. | |
|
Virchow R: Editoral Archiv fuer pathologische Anatomie und Physiologie und fuer klinische. Medizin. 8:231855. | |
|
Virchow R: Cellular Pathology. Hirschwald A: August Hirschwald; Berlin: 1858, PubMed/NCBI | |
|
Durante F: Nesso fisiopatologico tra la struttura dei nei materni e la genesi di alcuni tumori maligni. Arch Memorie ed Osservazioni di Chirurgia Pratica. 1874:217–226. 1874. | |
|
Cohnheim J: Congenitales, quergestreiftes muskelsarkon der nireren. Virchows Arch. 65:64–69. 1875. View Article : Google Scholar | |
|
Wilms M: Die Mischgeschwuelste. Leipzing; Arthur Georgi: 1899 | |
|
Ribbert H: Ueber Rückbildung an Zellen und Geweben und über die Entstehung der Geschwülste. Erwin Nägele; Stuttgart: 1897 | |
|
Ribbert, op. cit., Rückbildung (note 51). pp42–43, idem, op. cit., Beiträge (note 51). pp8–13, See also Johach, op. cit. (note 11). pp246–267 | |
|
Soto AM, Maffini MV and Sonnenschein C: Neoplasia as development gone awry: The role of endocrine disruptors. Int J Androl. 31:288–293. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Askanazy M: Die Teratome nach ihrem Bau, ihrem Verlauf, ihrer Genese und im Vergleich zum experimentellen Teratoid. Verhandl Deutsch Pathol. 11:39–82. 1907. | |
|
Stevens LC and Little CC: Spontaneous testicular teratomas in an inbred strain of mice. Proc Natl Acad Sci USA. 40:1080–1087. 1954. View Article : Google Scholar : PubMed/NCBI | |
|
Pierce GB and Dixon FJ: Testicular teratomas: I. The demonstration of teratogenesis by metamorphosis of multipotent cells. Cancer. 12:573–583. 1959. View Article : Google Scholar : PubMed/NCBI | |
|
Pierce GB and Verney EL: An in vitro and in vivo study of differentiation in teratocarcinomas. Cancer. 14:1017–1029. 1961. View Article : Google Scholar : PubMed/NCBI | |
|
Brinster RL: The effect of cells transferred into the mouse blastocyst on subsequent development. J Exp Med. 140:1049–1056. 1974. View Article : Google Scholar : PubMed/NCBI | |
|
Mintz B and Illmensee K: Normal genetically mosaic mice produced from malignant teratocarcinoma cells. Proc Natl Acad Sci USA. 72:3585–3589. 1975. View Article : Google Scholar : PubMed/NCBI | |
|
Grobstein C: The differentiation of such tissues may depend on inductive interactions between embryonic components. 13th Symposium of the Society for Development and Growth. Rudnick D: Princeton University Press; Princeton, NJ: pp. 233–256. 1954 | |
|
Rous P: A Sarcoma of the Fowl Transmissible by an Agent Separable from the Tumor Cells. J Exp Med. 13:397–411. 1911. View Article : Google Scholar : PubMed/NCBI | |
|
Macpherson I: Reversion in hamster cells transformed by Rous sarcoma virus. Science. 148:1731–1733. 1965. View Article : Google Scholar : PubMed/NCBI | |
|
Pollack RE, Green H and Todaro GJ: Growth control in cultured cells: Selection of sublines with increased sensitivity to contact inhibition and decreased tumor-producing ability. Proc Natl Acad Sci USA. 60:126–133. 1968. View Article : Google Scholar : PubMed/NCBI | |
|
Duran-Reynals F and Milford JJF: Growth of a chicken sarcoma virus in the chick embryo in the absence of neoplasia. Cancer Res. 3:578–584. 1943. | |
|
Dolberg DS and Bissell MJ: Inability of Rous sarcoma virus to cause sarcomas in the avian embryo. Nature. 309:552–556. 1984. View Article : Google Scholar : PubMed/NCBI | |
|
Braun AC: Bacterial and host factors concerned in determining tumor morphology in crown gall. Bot Gaz. 114:363–371. 1953. View Article : Google Scholar | |
|
Braun AC: A Demonstration of the recovery of the crown-gall tumor cell with the use of complex tumors of single-cell origin. Proc Natl Acad Sci USA. 45:932–938. 1959. View Article : Google Scholar : PubMed/NCBI | |
|
Rose SM: Epidermal dedifferentiation during blastema formation in regeneration limbs of Triturus viridescens. J Exp Zool. 108:337–362. 1948. View Article : Google Scholar : PubMed/NCBI | |
|
Wallingford HM: Transformations of renal tumors to normal tissue in regenerating limbs of salamanders. Science. 107:4571948.PubMed/NCBI | |
|
Gersch M: Zellentartung und Zellwucherung bei wirbellosen Tieren. Arch. Geschwulst-Forschung. 3:1–18. 1951. | |
|
Waddington CH: Cancer and the theory of organizers. Nature. 135:606–608. 1935. View Article : Google Scholar | |
|
Needham J: New advances in the chemistry and biology of organized growth. Proc R Soc London B Biol Sci. 29:1577–1626. 1936.PubMed/NCBI | |
|
Seilern-Aspang F and Kratochwil K: Induction and differentiation of an epithelial tumour in the newt (Triturus cristatus). J Embryol Exp Morphol. 10:337–356. 1962.PubMed/NCBI | |
|
McMichael H: Inhibition of growth of Shope rabbit papilloma by hypervitaminosis A. Cancer Res. 25:947–955. 1965.PubMed/NCBI | |
|
Saffiotti J, Montesano R, Sellakumar AR and Borg SA: Experimental cancer of the lung, inhibition by vitamin a of the induction of tracheobronchial squamous metaplasia and squamous cell tumors. Cancer. 20:857–864. 1967. View Article : Google Scholar : PubMed/NCBI | |
|
Davies RE: Effect of vitamin A on 7, 12-di-methylbenz(alpha) anthracene-induced papillomas in rhino mouse skin. Cancer Res. 27:237–241. 1967.PubMed/NCBI | |
|
Coleman WB, Wennerberg AE, Smith GJ and Grisham JW: Regulation of the differentiation of diploid and some aneuploid rat liver epithelial (stemlike) cells by the hepatic microenvironment. Am J Pathol. 142:1373–1382. 1993.PubMed/NCBI | |
|
Pierce GB: The cancer cell and its control by the embryo. Rous-Whipple Award lecture. Am J Pathol. 113:115–124. 1983. | |
|
Pierce GB, Lewis SH, Miller GJ, Moritz E and Miller P: Tumorigenicity of embryonal carcinoma as an assay to study control of malignancy by the murine blastocyst. Proc Natl Acad Sci USA. 76:6649–6651. 1979. View Article : Google Scholar : PubMed/NCBI | |
|
Pierce GB, Pantazis CG, Caldwell JE and Wells RS: Specificity of tumor formation by the blastocyst. Cancer Res. 42:1082–1087. 1982.PubMed/NCBI | |
|
Wells RS: An in vitro assay for regulation of embryonal carcinoma by the blastocyst. Cancer Res. 42:2736–2741. 1982.PubMed/NCBI | |
|
Podesta A, Beddington RSP, Wells RS and Pierce GB: The neurula stage mouse embryo in control of neuroblastoma. Proc Natl Acad Sci USA. 81:7608–7611. 1984. View Article : Google Scholar : PubMed/NCBI | |
|
Podesta AN, Mullins J, Pierce GB and Sells RS: The neurula state mouse embryos in control of neuroblastomas. Proc Natl Acad Sci USA. 81:7608–7611. 1984. View Article : Google Scholar : PubMed/NCBI | |
|
Gootwine E, Webb CG and Sachs L: Participation of myeloid leukaemia cells injected into embryos in haematopoietic differentiation in adult mice. Nature. 299:63–65. 1982. View Article : Google Scholar : PubMed/NCBI | |
|
Gerschenson M, Graves K, Carson SD, Wells RS and Pierce GB: Regulation of melanoma by the embryonic skin. Proc Natl Acad Sci USA. 83:7307–7310. 1986. View Article : Google Scholar : PubMed/NCBI | |
|
Pierce GB, Aguilar D, Hood G and Wells RS: Trophectoderm in control of murine embryonal carcinoma. Cancer Res. 44:3987–3996. 1984.PubMed/NCBI | |
|
DeCosse JJ, Gossens CL, Kuzma JF and Unsworth BR: Breast cancer: Induction of differentiation by embryonic tissue. Science. 181:1057–1058. 1973. View Article : Google Scholar : PubMed/NCBI | |
|
Biava PM, Fiorito A, Negro C and Mariani M: Effects of treatment with embryonic and uterine tissue homogenates on Lewis lung carcinoma development. Cancer Lett. 41:265–270. 1988. View Article : Google Scholar : PubMed/NCBI | |
|
Biava PM, Bonsignorio D and Hosha M: Cell proliferation curves of different human tumor lines after in vitro treatment with Zebrafish embryonic extracts. J Tumor Marker Oncol. 16:195–201. 2001. | |
|
Biava PM and Bonsignorio D: Cancer and cell differentiation: A model to explain malignancy. J Tumor Marker Oncol. 17:47–53. 2002. | |
|
Lee LM, Seftor EA, Bonde G, Cornell RA and Hendrix MJ: The fate of human malignant melanoma cells transplanted into zebrafish embryos: Assessment of migration and cell division in the absence of tumour formation. Dev Dyn. 233:1560–1570. 2005. View Article : Google Scholar : PubMed/NCBI | |
|
Cucina A, Biava PM, D'Anselmi F, Coluccia P, Conti F, di Clemente R, Miccheli A, Frati L, Gulino A and Bizzarri M: Zebrafish embryo proteins induce apoptosis in human colon cancer cells (Caco2). Apoptosis. 11:1617–1628. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Pierce GB and Johnson LD: Differentiation and cancer. In Vitro. 7:140–145. 1971. View Article : Google Scholar : PubMed/NCBI | |
|
Pierce GB and Wallace C: Differentiation of malignant to benign cells. Cancer Res. 31:127–134. 1971.PubMed/NCBI | |
|
Kenny PA and Bissell MJ: Tumor reversion: Correction of malignant behavior by microenvironmental cues. International J Cancer. 107:688–695. 2003. View Article : Google Scholar : PubMed/NCBI | |
|
Camacho LH: Clinical application of retinoids in cancer medicine. J Biol Regul Homeost Agents. 17:98–114. 2003.PubMed/NCBI | |
|
Pitha-Rowe I, Petty WJ, Kitareewan S and Dmitrovsky E: Retinoid target genes in acute promyelocytic leukemia. Leukemia. 17:1723–1730. 2003. View Article : Google Scholar : PubMed/NCBI | |
|
Segalla S, Rinaldi L, Kilstrup-Nielsen C, Badaracco G, Minucci S, Pelicci PG and Landsberger N: Retinoic acid receptor alpha fusion to PML affects in transcriptional and chromatin-remodeling properties. Mol Cell Biol. 23:8795–808. 2003. View Article : Google Scholar : PubMed/NCBI | |
|
Alcalay M, Meani N, Gelmetti V, Fantozzi A, Fagioli M, Orleth A, Riganelli D, Sebastiani C, Cappelli E, Casciari C, et al: Acute myeloid leukemia fusion proteins deregulate genes involved in stem cell maintenance and DNA repair. J Clin Invest. 112:1751–1761. 2003. View Article : Google Scholar : PubMed/NCBI | |
|
Strickland S and Madavi V: The induction of differentiation in teratocarcinoma stem cells by retinoic acid. Cell. 15:393–403. 1978. View Article : Google Scholar : PubMed/NCBI | |
|
Trump DL: Retinoids in bladder, testes and prostate cancer: Epidemiologic, preclinical and clinical observations. Leukemia. 8 (Suppl 3):S50–S54. 1994.PubMed/NCBI | |
|
Breitman TR, Selonick SE and Collins SJ: Induction of differentiation of the human promyelocytic leukemia cell line (HL-60) by retinoic acid. Proc Natl Acad Sci USA. 77:2936–2940. 1980. View Article : Google Scholar : PubMed/NCBI | |
|
Huang ME, Ye YC, Chen SR, Chai JR, Lu JX, Zhoa L, Gu LJ and Wang ZY: Use of all-trans retinoic acid in the treatment of acute promyelocytic leukemia. Blood. 72:567–572. 1988. View Article : Google Scholar : PubMed/NCBI | |
|
Dragnev KH, Petty WJ and Dmitrovsky E: Retinoid targets in cancer therapy and chemoprevention. Cancer Biol Ther. 2 (Suppl 1):S150–S156. 2003. View Article : Google Scholar : PubMed/NCBI | |
|
Melnick A and Licht JD: Deconstruction a disease: RARalpha, its fusion partners, and their roles in the pathogenesis of acute promyelocytic leukemia. Blood. 99:3167–3215. 1999. View Article : Google Scholar : PubMed/NCBI | |
|
Warrell RP Jr, Frankel SR, Miller WH Jr, Scheinberg DA, Itri LM, Hittelman WN, Vyas R, Andreeff M, Tafuri A and Jakubowski A: Differention therapy of acute promyelocytic leukemia with tretinoin (all-trans-retinoic acid). N Engl J Med. 324:1385–1393. 1991. View Article : Google Scholar : PubMed/NCBI | |
|
Vogelstein B, Papadopoulos N, Velculescu VE, Zhou S, Diaz LA and Kinzler KW: Cancer genome landscapes. Science. 339:1546–1558. 6127. View Article : Google Scholar : PubMed/NCBI | |
|
Bertolaso M: Philosophy of Cancer-A Dynamic and Relational View. Springer; New York, NY: 2016, PubMed/NCBI | |
|
Rohdenburg GL: Fluctuations in the growth of malignant tumors in man, with especial reference to spontaneous regression. J Cancer Res. 3:192–221. 1918. | |
|
Cushing H and Wollbach S: The transformation of malignant paravertebral Sympathicoblastoma into a benign ganglioneuroma. Am J Pathol. 3:203–216.7. 1927.PubMed/NCBI | |
|
Bumpus HC: The apparent disappearance of pulmonary metastasis in a case of hypernephroma following nephrectomy. J Urol. 20:185–191. 1927. View Article : Google Scholar | |
|
Everson TC and Cole WH: Spontaneous Regression of Cancer. W.B Saunders; Philadelphia, PA: 1966, PubMed/NCBI | |
|
Cole WH: Spontaneous regression of cancer and the importance of finding its cause. Nat Cancer Inst Mnogr. 44:5–9. 1976.PubMed/NCBI | |
|
Challis GB and Stam HJ: The spontaneous regression of cancer. A review of cases from 1900 to 1987. Acta Oncol. 29:545–549. 1990. View Article : Google Scholar : PubMed/NCBI | |
|
O'Regan B and Hirschberg C: Spontaneous Regression. An Annotated Bibliography Sausalito CA: Institute of Noetic Science; 1993 | |
|
Papac RJ: Spontaneous regression of cancer: Possible mechanisms. In Vivo. 12:571–578. 1998.PubMed/NCBI | |
|
Livraghi T, Meloni F, Frosi A, Lazzaroni S, Bizzarri TM, Frati L and Biava PM: Treatment with stem cell differentiation stage factors of intermediate-advanced hepatocellular carcinoma: An open randomized clinical trial. Oncol Res. 15:399–408. 2005. View Article : Google Scholar : PubMed/NCBI | |
|
Telerman A, Tuynder M, Dupressoir T, Robaye B, Sigaux F, Shaulian E, Oren M, Rommelaere J and Amson R: A model for tumor suppression using H-1 parvovirus. Proc Natl Acad Sci USA. 90:8702–8706. 1993. View Article : Google Scholar : PubMed/NCBI | |
|
Tuynder M, Susini L, Prieur S, Besse S, Fiucci G, Amson R and Telerman A: Biological models and genes of tumor reversion: Cellular reprogramming through tpt1/TCTP and SIAH-1. Proc Natl Acad Sci USA. 99:14976–1481. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Telerman A and Amson R: The molecular programme of tumour reversion: The steps beyond malignant transformation. Nat Rev Cancer. 9:206–216. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Tuynder M, Fiucci G, Prieur S, Lespagnol A, Géant A, Beaucourt S, Duflaut D, Besse S, Susini L, Cavarelli J, et al: Translationally controlled tumor protein is a target of tumor reversion. Proc Natl Acad Sci USA. 101:15364–15369. 2004. View Article : Google Scholar : PubMed/NCBI | |
|
Thaw P, Baxter NJ, Hounslow AM, Price C, Waltho JP and Craven CJ: Structure of TCTP reveals unexpected relationship with guanine nucleotide-free chaperones. Nat Struct Biol. 8:701–704. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Proietti S, Cucina A, Pensotti A, Biava PM, Minini M, Monti N, Catizone A, Ricci G, Leonetti E, Harrath AH, et al: Active fraction from embryo fish extracts induces reversion of the malignant invasive phenotype in breast cancer through down-regulation of TCTP and modulation of E-cadherin/β-catenin pathway. Int J Mol Sci. 20:21512019. View Article : Google Scholar : PubMed/NCBI | |
|
Weaver VM, Petersen OW, Wang F, Larabell CA, Briand P, Damsky C and Bissell MJ: Reversion of the malignant phenotype of human breast cells in three-dimensional culture and in vivo by integrin blocking antibodies. J Cell Biol. 137:231–245. 1997. View Article : Google Scholar : PubMed/NCBI | |
|
Hendrix MJ, Seftor EA, Seftor RE, Kasemeier-Kulesa J, Kulesa PM and Postovit LM: Reprogramming metastatic tumour cells with embryonic microenvironments. Nat Rev Cancer. 7:246–255. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Tabibzadeh S and Hemmati-Brivanlou A: Lefty at the crossroads of ‘stemness’ and differentiative events. Stem Cells. 24:1998–2006. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Postovit LM, Margaryan NV, Seftor EA, Kirschmann DA, Lipavsky A, Wheaton WW, Abbott DE, Seftor RE and Hendrix MJ: Human embryonic stem cell microenvironment suppresses the tumorigenic phenotype of aggressive cancer cells. Proc Natl Acad Sci USA. 105:4329–4334. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Topczewska JM, Postovit LM, Margaryan NV, Sam A, Hess AR, Wheaton WW, Nickoloff BJ, Topczewski J and Hendrix MJ: Embryonic and tumorigenic pathways converge via Nodal signaling: Role in melanoma aggressiveness. Nat Med. 12:925–932. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Costa FF: Non-coding RNAs: Lost in translation? Gene. 386:1–10. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Garzon R, Fabbri M, Cimmino A, Calin GA and Croce CM: MicroRNA expression and function in cancer. Trends Mol Med. 12:580–587. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Costa FF, Seftor EA, Bischof JM, Kirschmann DA, Strizzi L, Arndt K, Bonaldo Mde F, Soares MB and Hendrix MJ: Epigenetically reprogramming metastatic tumor cells with an embryonic microenvironment. Epigenomics. 1:387–398. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Card DA, Hebbar PB, Li L, Trotter KW, Komatsu Y, Mishina Y and Archer TK: Oct4/Sox2-regulated miR-302 targets cyclin D1 in human embryonic stem cells. Mol Cell Biol. 28:6426–6438. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Krebs LT, Iwai N, Nonaka S, Welsh IC, Lan Y, Jiang R, Saijoh Y, O'Brien TP, Hamada H and Gridley T: Notch signaling regulates left-right asymmetry determination by inducing Nodal expression. Genes Dev. 17:1207–1212. 2003. View Article : Google Scholar : PubMed/NCBI | |
|
Morgan DO: Cyclin-dependent kinases: Engines, clocks, and microprocessors. Annu Rev Cell Dev Biol. 13:261–291. 1997. View Article : Google Scholar : PubMed/NCBI | |
|
Giuffrida D, Rogers IM, Nagy A, Calogero AE, Brown TJ and Casper RF: Human embryonic stem cells secrete soluble factors that inhibit cancer cell growth. Cell Prolif. 42:788–798. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Novak P, Jensen TJ, Garbe JC, Stampfer MR and Futscher BW: Stepwise DNA methylation changes are linked to escape from defined proliferation barriers and mammary epithelial cell immortalization. Cancer Res. 69:5251–5258. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Hinshelwood RA and Clark SJ: Breast cancer epigenetics: Normal human mammary epithelial cells as a model system. J Mol Med. 86:1315–1328. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Allegrucci C, Rushton MD, Dixon JE, Sottile V, Shah M, Kumari R, Watson S, Alberio R and Johnson AD: Epigenetic reprogramming of breast cancer cells with oocyte extracts. Mol Cancer. 10:72011. View Article : Google Scholar : PubMed/NCBI | |
|
Saad N, Alberio R, Johnson AD, Emes RD, Giles TC, Clarke P, Grabowska AM and Allegrucci C: Cancer reversion with oocyte extracts is mediated by cell cycle arrest and induction of tumour dormancy. Oncotarget. 9:16008–16027. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Tripathi A, Kashyap A, Tripathi G, Yadav J, Bibban R, Aggarwal N, Thakur K, Chhokar A, Jadli M, Sah AK, et al: Tumor reversion: A dream or a reality. Biomark Res. 9:312021. View Article : Google Scholar : PubMed/NCBI | |
|
Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K and Yamanaka S: Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 131:861–872. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Feinberg AP: Phenotypic plasticity and the epigenetics of human disease. Nature. 447:433–440. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Redmer T, Diecke S, Grigoryan T, Quiroga-Negreira A, Birchmeier W and Besser D: E-cadherin is crucial for embryonic stem cell pluripotency and can replace OCT4 during somatic cell reprogramming. EMBO Rep. 12:720–726. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Feng B, Ng JH, Heng JC and Ng HH: Molecules that promote or enhance reprogramming of somatic cells to induced pluripotent stem cells. Cell Stem Cell. 4:301–312. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Smith ZD, Sindhu C and Meissner A: Molecular features of cellular reprogramming and development. Nat Rev Mol Cell Biol. 17:139–154. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Yoo J and Kim J, Baek S, Park Y, Im H and Kim J: Cell reprogramming into the pluripotent state using graphene based substrates. Biomaterials. 35:8321–8329. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Bizzarri M, Palombo A and Cucina A: Theoretical aspects of systems biology. Prog Biophys Mol Biol. 112:33–43. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Nieto MA, Huang RY, Jackson RA and Thiery JP: EMT: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Abad M, Mosteiro L, Pantoja C, Cañamero M, Rayon T, Ors I, Graña O, Megías D, Domínguez O, Martínez D, et al: Reprogramming in vivo produces teratomas and iPS cells with totipotency features. Nature. 502:340–345. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Ohnishi K, Semi K, Yamamoto T, Shimizu M, Tanaka A, Mitsunaga K, Okita K, Osafune K, Arioka Y, Maeda T, et al: Premature termination of reprogramming in vivo leads to cancer development through altered epigenetic regulation. Cell. 156:663–677. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Inman JL, Robertson C, Mott JD and Bissell MJ: Mammary gland development: Cell fate specification, stem cells and the microenvironment. Development. 142:1028–1042. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Bizzarri M and Giuliani A: Representing cancer cell trajectories in a phase-space diagram: Switching cellular states by biological phase transitions. Applied Statistics for Network Biology: Methods in Systems Biology. Dehmer M, Emmert-Streib F, Graber A and Salvador A: Wiley-VCH Verlag GmbH & Co.; pp. 377–403. 2011, View Article : Google Scholar | |
|
Lin SL, Chang DC, Chang-Lin S, Lin CH, Wu DT, Chen DT and Ying SY: Mir-302 reprograms human skin cancer cells into a pluripotent ES-cell-like state. RNA. 14:2115–2124. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Utikal J, Maherali N, Kulalert W and Hochedlinger K: Sox2 is dispensable for the reprogramming of melanocytes and melanoma cells into induced pluripotent stem cells. J Cell Sci. 122:3502–3510. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Rapino F, Robles EF, Richter-Larrea JA, Kallin EM, Martinez-Climent JA and Graf T: C/EBPα induces highly efficient macrophage transdifferentiation of B lymphoma and leukemia cell lines and impairs their tumorigenicity. Cell Rep. 3:1153–1163. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Huang P, Zhang L, Gao Y, He Z, Yao D, Wu Z, Cen J, Chen X, Liu C, Hu Y, et al: Direct reprogramming of human fibroblasts to functional and expandable hepatocytes. Cell Stem Cell. 14:370–384. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
McClellan JS, Dove C, Gentles AJ, Ryan CE and Majeti R: Reprogramming of primary human Philadelphia chromosome-positive B cell acute lymphoblastic leukemia cells into nonleukemic macrophages. Proc Natl Acad Sci USA. 112:4074–4079. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Zhou S, Abdouh M, Arena V, Arena M and Arena GO: Reprogramming malignant cancer cells toward a benign phenotype following expo-sure to human embryonic stem cell microenvironment. PLoS One. 12:e01698992017. View Article : Google Scholar : PubMed/NCBI | |
|
Ishay-Ronen D, Diepenbruck M, Kalathur RKR, Sugiyama N, Tiede S, Ivanek R, Bantug G, Morini MF, Wang J, Hess C and Christofori G: Gain fat-lose metastasis: Converting invasive breast cancer cells into adipocytes inhibits cancer metastasis. Cancer Cell. 35:17–32.e6. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Cheng Z, He Z, Cai Y, Zhang C, Fu G, Li H, Sun W, Liu C, Cui X, Ning B, et al: Conversion of hepatoma cells to hepatocyte-like cells by defined hepatocyte nuclear factors. Cell Res. 29:124–135. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Li Y, Agrawal I and Gong Z: Reversion of tumor hepatocytes to normal hepatocytes during liver tumor regression in an oncogene-expressing transgenic zebrafish model. Dis Model Mech. 12:dmm0395782019. View Article : Google Scholar : PubMed/NCBI | |
|
Pensotti A, Bertolaso M and Bizzarri M: Is cancer reversible? Rethinking carcinogenesis models-a new epistemological tool. Biomolecules. 13:7332023. View Article : Google Scholar : PubMed/NCBI | |
|
Longo G, Miquel PA, Sonnenschein C and Soto AM: Is information a proper observable for biological organization? Prog Biophys Mol Biol. 109:108–114. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Kholodenko BN, Kolch W and Rukhlenko OS: Reversing pathological cell states: The road less travelled can extend the therapeutic horizon. Trends Cell Biol. 33:913–923. 2023. View Article : Google Scholar : PubMed/NCBI |









