


Resveratrol inhibits the proliferation of human melanoma cells by inducing G1/S cell cycle arrest and apoptosis
- Authors:
- Published online on: October 21, 2014 https://doi.org/10.3892/mmr.2014.2716
- Pages: 400-404
Abstract
Introduction
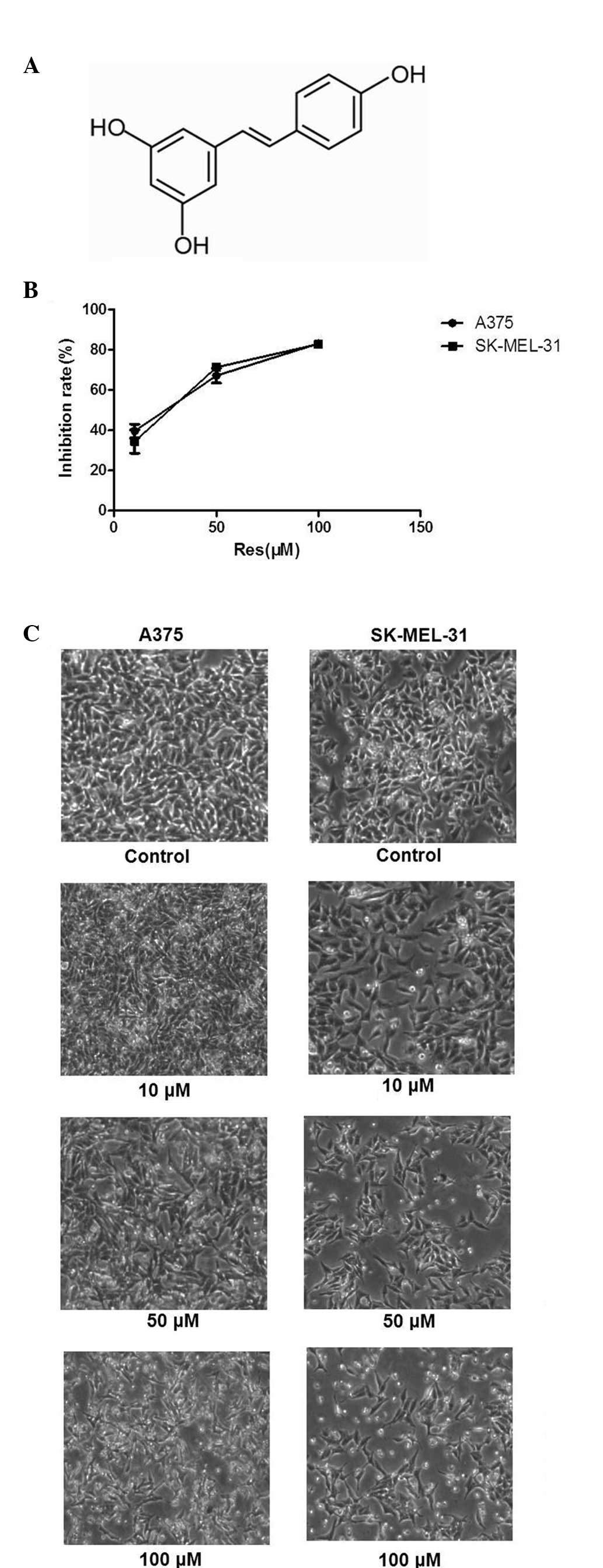
Resveratrol (Res; C14H12O3; PubChem, CID: 445154; Fig. 1A) is primarily found in the skin of grapes and has been demonstrated to exhibit health-promoting benefits to the coronary, neurological, hepatic and cardiovascular systems (1–3). In addition, Res inhibits the proliferation of tumor cells of different etiologies (4,5). The biological properties of Res have been described in detail (6–9). The predominant form of resveratrol in plants is the glycosylated (3-O-β-D-glucosides) form (also termed the piceid form). It has traditionally been used as an anti-inflammatory agent. Other beneficial properties of resveratrol include antioxidant effects, cardioprotection and increased longevity. The antitumor activities of Res are mediated through several cell signaling pathways, including cell cycle arrest, suppression of tumor cell proliferation, induction of apoptosis and differentiation, angiogenesis and the inhibition of invasion, adhesion and metastasis (10). Skin cancer is one of the major causes of cancer-associated mortality worldwide. In addition, human cutaneous malignant melanoma is an aggressive cause of mortality, which exhibits a rising trend every year (9). In order to inhibit the development of cancer, pharmacological or natural chemopreventive and chemotherapeutic agents are commonly used (12).
p53 is a frequent target for mutation in various types of human tumor. It functions as a cell nucleus phosphate protein, which responds to various types and levels of stress arising from apoptosis, cell cycle arrest, senescence, DNA repair and cell metabolism (13–15). A previous study demonstrated that gambogic acid, an efficient apoptosis inducer, was able to repress the expression of B-cell lymphoma 2 (Bcl-2) via increasing the level of p53 in MCF-7 cells (16). Although previous studies have described intracellular changes leading to cell cycle arrest or apoptosis in response to Res treatment, the precise mechanisms underlying Res-regulated tumor growth remain to be fully elucidated (20–22). In the present study, the anti-cancer effects of Res in human melanoma A375 and SK-MEL-31 cells were assessed. The results demonstrated for the first time, to the best of our knowledge, that the regulation of caspase and p53 proteins was involved in the induction of apoptosis and cell cycle arrest by Res.
Materials and methods
Cell culture
Human melanoma A375 and SK-MEL-31 cells, obtained from the Affiliated Hospital of Guangdong Medical College (Guangdong, China), were grown as a suspension culture in Dulbecco’s modified Eagle’s medium (Invitrogen Life Technologies, Carlsbad, CA, USA) supplemented with 10% (v/v) fetal bovine serum (Gibco-BRL, Carlsbad, CA, USA) 100 U/ml penicillin and 100 U/ml streptomycin. They were maintained in a humidified atmosphere of 95% air and 5% CO2 at 37°C.
Reagents
Res was purchased from Sigma-Aldrich (St. Louis, MO, USA). Monoclonal human anti-rabbit antibodies to caspase-9, caspase-3, Bcl-2, Bcl-2-associated X protein (Bax), p53 and GAPDH were obtained from Cell Signaling Technology, Inc. (Beverly, MA, USA) and goat anti-rabbit IgG-horseradish peroxidase (EarthOx Life Sciences, Millbrae, CA, USA) was used as a secondary antibody.
Cell viability assays
The cell densities were adjusted to 2×104 cells/100 μl. The cells were seeded into a 96-well plate and treated with 10, 50 and 100 μM Res for 24 h. The cell viability was then assessed using a Cell Titer 96 AQueous One Solution Cell Proliferation Assay kit (Promega Corporation, Madison, WI, USA) according to the manufacturer’s instructions.
Analysis of cell apoptosis and cell cycle arrest
Cells were pretreated with 10, 50 and 100 μM Res for 48 h. Cells were quantified using a Cell Cycle Analysis kit (Beyotime Institute of Biotechnology) and a PE Annexin V Apoptosis Detection kit I (BD Biosciences, Franklin Lakes, NJ, USA), respectively. Analysis was preformed using flow cytometry (FACSCalibur; Beckton Dickinson, Franklin Lakes, NJ, USA) and then analyzed using ModFit and CellQuest software (BD Biosciences).
Western blot analysis
Cells were lysed in lysis buffer (100 mM Tris-hydrochloride, 4% pH 6.8 (m/v) sodium dodecyl sulfonate, 20% (v/v), glycerol, 200 mM mercaptoethanol, 1 mM phenylmethyl sulfonylfluoride and 1 g/ml aprotinin). The total protein concentration in the supernatants was detected using a BCA Protein assay kit (Beyotime Institute of Biotechnology). Proteins were then transferred onto nitrocellulose membranes (Millipore, Billerica, MA, USA). Detection was performed using an Odyssey Infrared Imaging System (LI-COR Biosciences, Lincoln, NE, USA).
Statistical analysis
The data were analyzed using GraphPad Prism 5 software (GraphPad Software, Inc., San Diego, CA, USA). All results are expressed as the mean ± standard deviation from triplicate experiments performed in a parallel manner, unless otherwise indicated.
Results
Res induces growth inhibition, proliferation and cell apoptosis in A375 and SK-MEL-31 cells
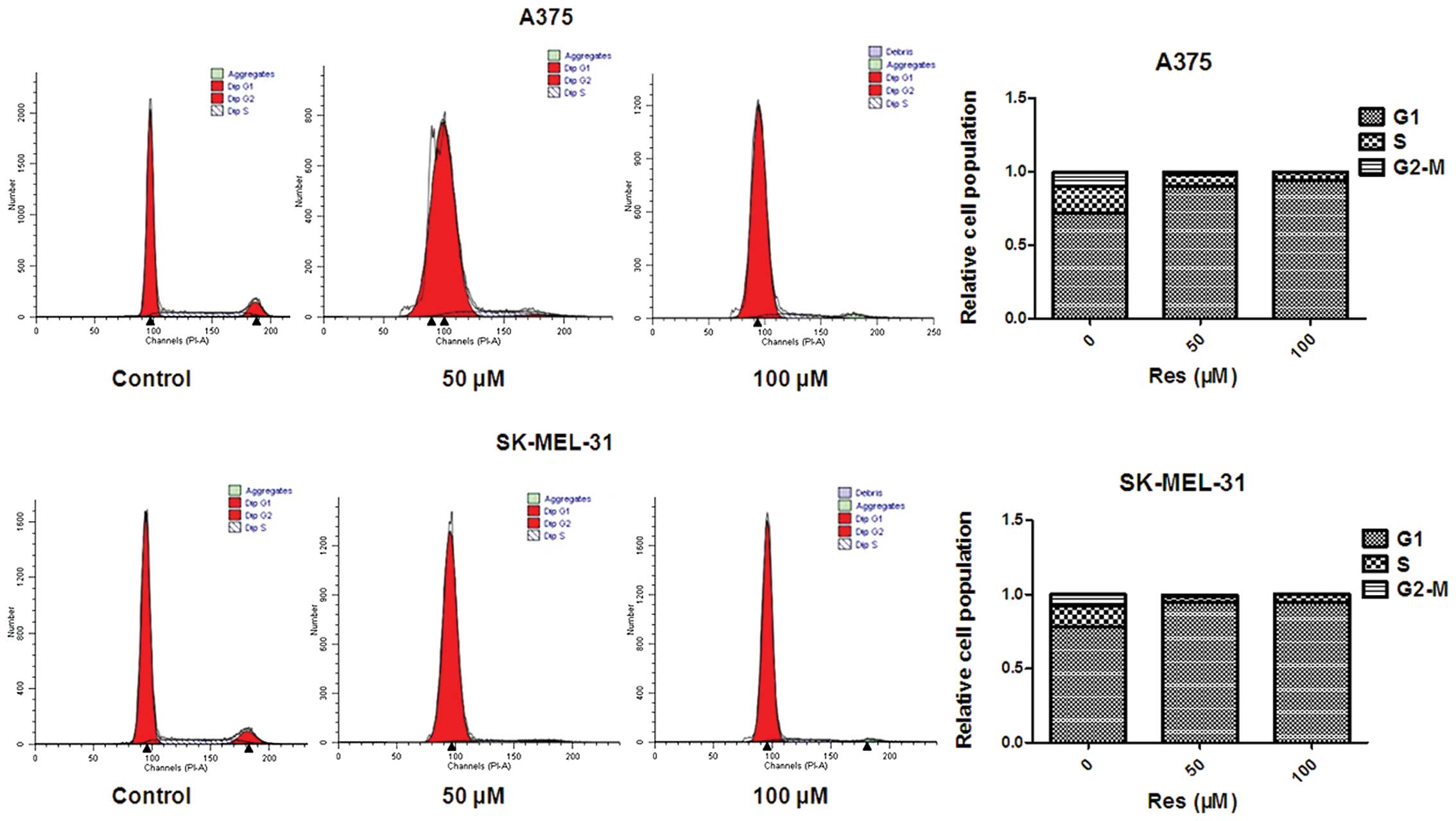
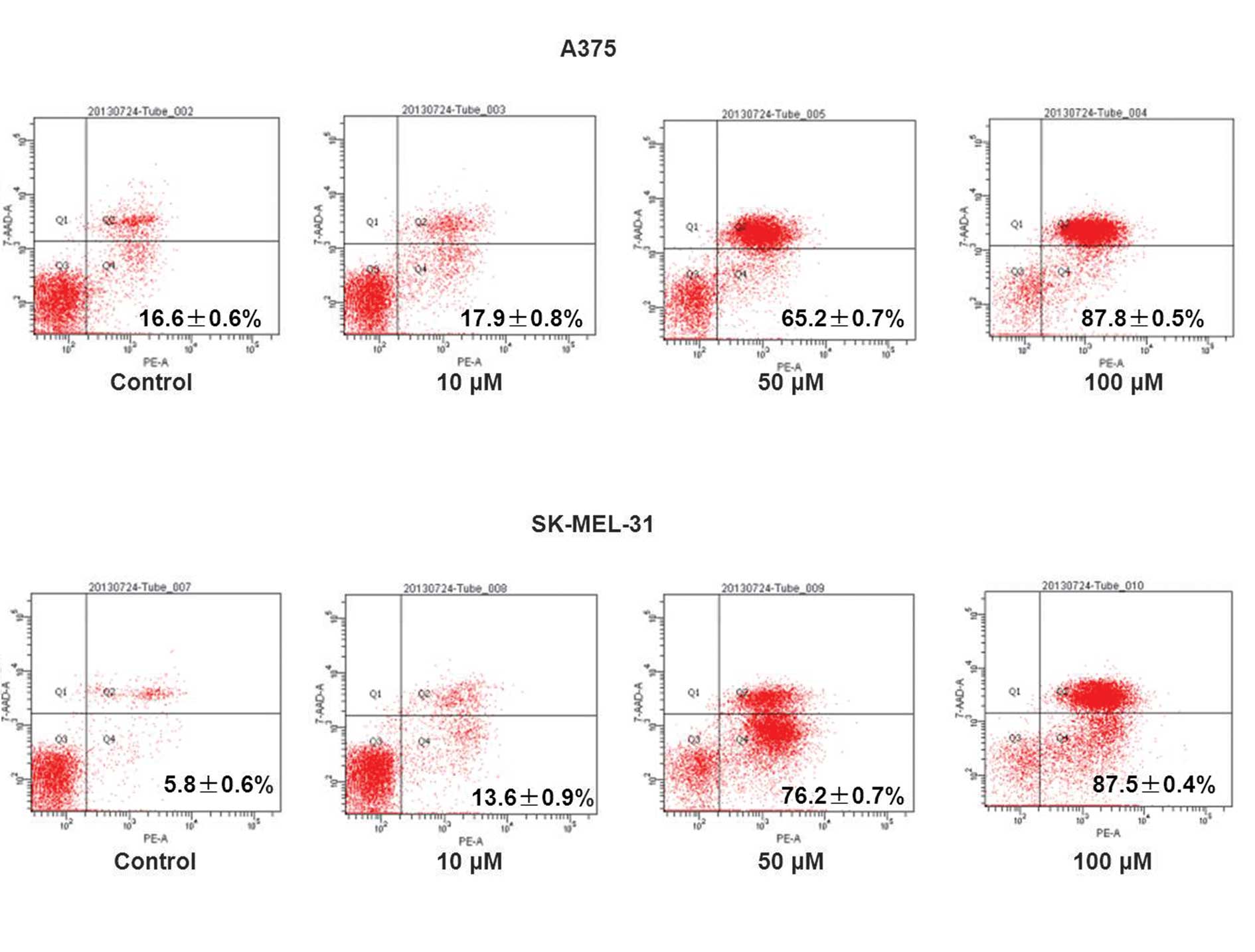
In the Res-treated cells, certain cells became round and floating and a marked reduction in cell viability was observed over the experimental period in a concentration-dependent manner (Fig. 1B and C). Following treatment with 50 and 100 μM Res, inhibition of the cell cycle was observed in the A375 and SK-MEL-31 cells (Fig. 2). The population of cells in the G1 phase increased in A375 (78–93%) and SK-MEL-31 (78–94%) cells, coinciding with a reduction in cells in the S phase, when compared with the vehicle-treated control. This result demonstrated that the primary growth inhibitory effect of Res was due to inhibition of the cell cycle at the G1 phase. Cell apoptosis was detected using flow cytometry. Data revealed that Res induced cell apoptosis in a concentration-dependent manner (Fig. 3). In addition, the Q2 and Q4 cell population in A375 (16.6–87.8%) and SK-MEL-31 (5.8–87.5%) cells increased compared with the vehicle-treated control (Fig. 3).
Res induces the expression of cell cycle and apoptotic-related proteins in A375 and SK-MEL-31 cells
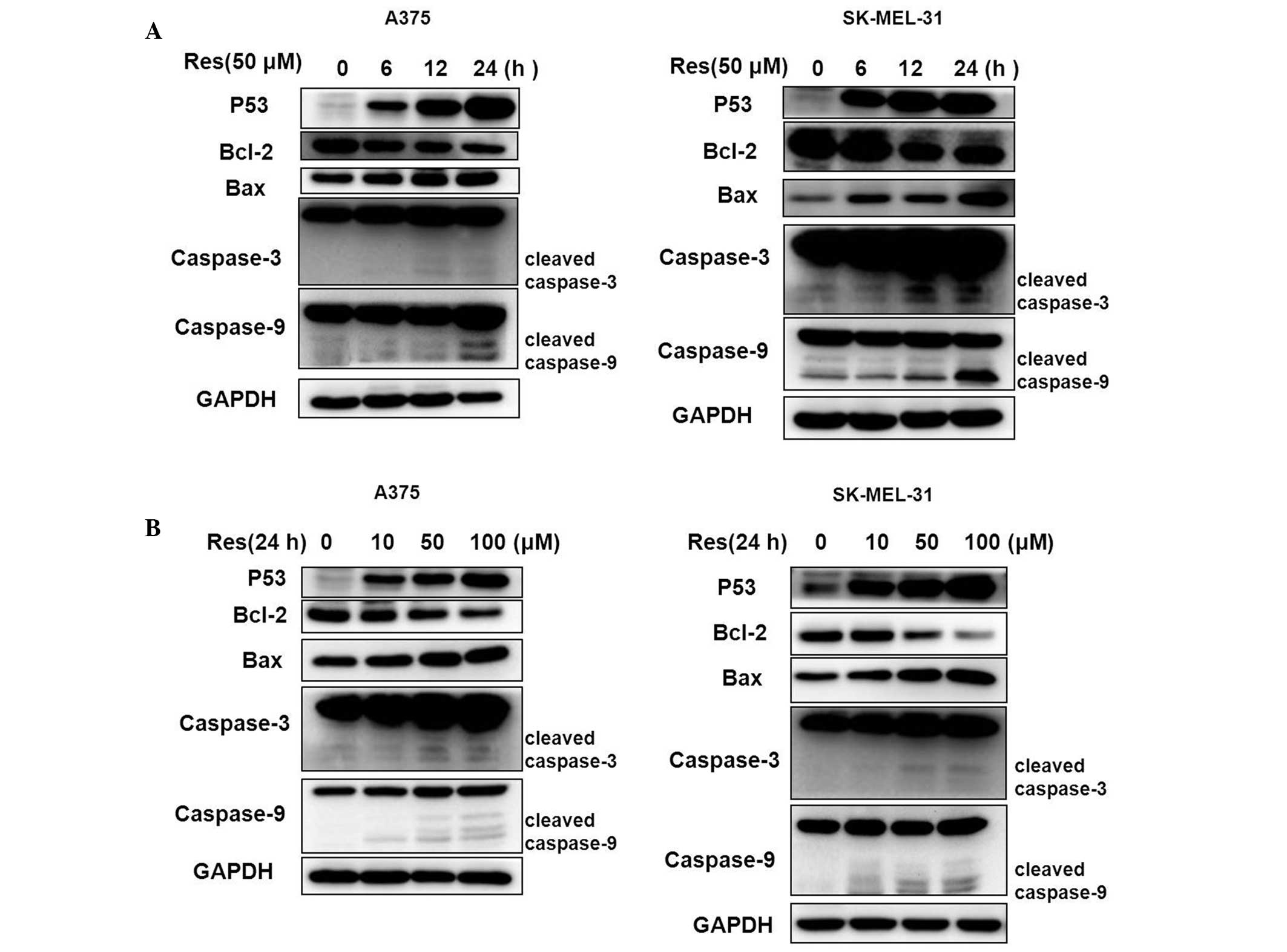
Western blot analysis demonstrated that cell cycle and apoptotic-related proteins altered following treatment with Res (Fig. 4). The Bcl-2 protein was downregulated, however, the apoptotic proteins of p53, Bax, caspase-3 and caspase-9 were markedly upregulated in a concentration- and time-dependent manner.
Discussion
Res, a natural plant polyphenol compound, has been extensively investigated for several years due to its various potential health-promoting benefits (8,20–23). In the present study, the effects of Res in human melanoma A375 and SK-MEL-31 cell lines was investigated. The inhibitory and apoptotic-promoting effects of Res on the growth of two human melanoma cells were determined, these cells have different genetic aberrations and acquired growth aggressiveness. The results from the cell viability and apoptotic rate assay demonstrated that the anticancer properties of Res in A375 and SK-MEL-31 cells were almost identical. The cell viability, G1 phase cell-cycle arrest and apoptotic rates increased in a concentration-dependent manner. Subsequently, the cell cycle and apoptotic-related proteins p53, Bcl-2, Bax, caspase-3 and caspase-9 were analyzed. The protein expression of Bcl-2 reduced, however, the protein expression of p53, Bax, caspase-3 and caspase-9 were markedly enhanced compared with the control. Cleaved caspase-9, cleaved caspase-3 and the ratio of Bax and Bcl-2 increased gradually in a concentration- and time-dependent manner. Caspase-9 and caspase-3 activation is a crucial step in apoptotic cell death (24,25) and the increased expression of cleaved caspase-9 and cleaved caspase-3 may be considered as a marker of apoptosis. The apoptosis-inducing effect is more dependent on the balance of Bcl-2 and Bax than on the quantity of Bcl-2 alone, which is important in cell proliferation. The steady state of cell survival is decided by the balance of Bcl-2 and Bax expression (26,27). Previous studies have demonstrated that the apoptotic inducer, p53, induces cell growth arrest, apoptosis and senescence in response to different stimuli and this was associated with cancer cell metastasis (28–31). The increased expression level of the Bax/Bcl-2 protein ratio in cells treated with Res suggested that the p53 and Bax/Bcl-2 proteins are important in Res-induced apoptosis.
Acknowledgements
This study was supported, in part, by the following grants: The National Natural Science Fund (no. 81041099) and the Guangdong Province Natural Science Fund (no. S2011010003750), China. This study was also supported by the Institute of Neurology, Affiliated Hospital of Guangdong Medical College and National Natural Science Foundation of China (no. 81102066), the Natural Science Foundation of Guangdong Province of China (no. S2011040003129) and the Medical Research Foundation of Guangdong Province of China (no. A2011441).
References
|
Chan V, Fenning A, Iver A, et al: Resveratol improves cardiovascular function in DOCA-salt hypertensive rats. Curr Pharm Biotechnol. 12:429–436. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Jiang L, Gu Y, Ye J, Liu F, et al: Resveratrol prevents hepatic steatosis induced by hepatitis C virus core protein. Biotechnol Lett. 34:2205–2212. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Zhang F, Liu J and Shi JS: Anti-inflammatory activities of resveratrol in the brain: role of resveratrol in microglial activation. Eur J Pharmacol. 25:1–7. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Lin JN, Lin VC, Rau KM, et al: Resveratrol modulates tumor cell proliferation and protein translation via SIRT1-dependent AMPK activation. J Agric Food Chem. 58:1584–92. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Huang F, Wu XN, Chen J, et al: Resveratrol reverses multidrug resistance in human breast cancer doxorubicin-resistant cells. Exp Ther Med. 7:1611–1616. 2014.PubMed/NCBI | |
|
Fulda S: Resveratrol and derivatives for the prevention and treatment of cancer. Drug Discov Today. 15:757–765. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Calabrese EJ, Mattson MP and Calabrese V: Dose response biology: the case of resveratrol. Hum Exp Toxicol. 29:1034–1037. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Park SJ, Ahmad F, Philp A, et al: Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell. 148:421–433. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Bishayee A, Politis T and Darvesh AS: Resveratrol in the chemoprevention and treatment of hepatocellular carcinoma. Cancer Treat Rev. 36:43–53. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Bishayee A: Cancer prevention and treatment with resveratrol: from rodent studies to clinical trials. Cancer Prev Res (Philia). 2:409–418. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Hsiao YP, Yu CS, Yu CC, et al: Triggering apoptotic death of human malignant melanoma a375.s2 cells by bufalin: Involvement of caspase cascade-dependent and independent mitochondrial signaling pathways. Evi Based Complemen Alternat Med. 2012:5912412012. | |
|
Kuo JH, Chu YL, Yang JS, et al: Cantharidin induces apoptosis in human bladder cancer TSGH 8301 cells through mitochondria-dependent signal pathways. Int J Oncol. 37:1243–1250. 2010.PubMed/NCBI | |
|
Vazquez A, Bond EE, Levine AJ and Bond GL: The genetics of the p53 pathway, apoptosis and cancer therapy. Nat Rev Drug Discov. 7:979–987. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Brady CA, Jiang D, Mello SS, et al: Distinct p53 transcriptional programs dictate acute DNA-damage responses and tumor suppression. Cell. 145:571–583. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Chee JL, Saidin S, Lane DP, et al: Wild-type and mutant p53 mediate cisplatin resistance through interaction and inhibition of active caspase-9. Cell Cycle. 12:278–288. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Gu H, Rao S, Zhao J, et al: Gambogic acid reduced bcl-2 expression via p53 in human breast MCF-7 cancer cells. J Cancer Res Clin Oncol. 135:1777–1782. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Liu B, Zhou Z, Zhou W, et al: Resveratrol inhibits proliferation in human colorectal carcinoma cells by inducing G1/S-phase cell cycle arrest and apoptosis through caspase/cyclin-CDK pathways. Mol Med Rep. 10:1697–1702. 2014.PubMed/NCBI | |
|
Li P, Yang S, Dou M, et al: Synergic effects of artemisinin and resveratrol in cancer cells. J Cancer Res Clin Oncol. 2014:Epub ahead of print. 2014. | |
|
Zhang P, Li H, Yang B, et al: Biological significance and therapeutic implication of resveratrol-inhibited Wnt, Notch and STAT3 signaling in cervical cancer cells. Genes Cancer. 5:154–64. 2014.PubMed/NCBI | |
|
Szekeres T, Saiko P, Fritzer-Szekeres M, Djavan B and Jäger W: Chemopreventive effects of resveratrol and resveratrol derivatives. Ann NY Acad Sci. 1215:89–95. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Nutakul W, Sobers HS, Qiu P, et al: Inhibitory effects of resveratrol and pterostilbene on human colon cancer cells: a side-by-side comparison. J Agric Food Chem. 59:10964–10970. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Piotrowska H, Kucinska M and Murias M: Biological activity of piceatannol: Leaving the shadow of resveratrol. Mutat Res. 750:60–82. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Um JH, Park SJ, Kang H, et al: AMP-activated protein kinase-deficient mice are resistant to the metabolic effects of resveratrol. Diabetes. 59:554–563. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Zhang Y, Liu H, Jin J, Zhu X, Lu L and Jiang H: The role of endogenous reactive oxygen species in oxymatrine-induced caspase-3-dependent apoptosis in human melanoma A375 cells. Anticancer Drugs. 21:494–501. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Miyata M, Kambe M, Tajima O, et al: Membrane sialidase NEU3 is highly expressed in human melanoma cells promoting cell growth with minimal changes in the composition of gangliosides. Cancer Sci. 102:2139–2149. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Tasyriq M, Najmuldeen IA, In LL, Mohamad K, Awang K and Hasima N: 7 alpha-Hydroxy-beta-Sitosterol from Chisocheton tomentosus induces apoptosis via dysregulation of cellular Bax/Bcl-2 ratio and cell cycle arrest by downregulating ERK1/2 activation. Evid Based Complement Alternat Med. 2012:7653162012.PubMed/NCBI | |
|
Wu S, Liu B, Zhang Q, et al: Dihydromyricetin reduced Bcl-2 expression via p53 in human hepatoma HepG2 cells. PloS One. 8:e768862013. View Article : Google Scholar : PubMed/NCBI | |
|
Zhu HB, Yang K, Xie YQ, Lin YW, Mao QQ and Xie LP: Silencing of mutant p53 by siRNA induces cell cycle arrest and apoptosis in human bladder cancer cells. World J Surg Oncology. 11:222013. View Article : Google Scholar : PubMed/NCBI | |
|
Yeudall WA, Wrighton KH and Deb S: Mutant p53 in cell adhesion and motility. Methods Mol Biol. 962:135–146. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Voskamp P, Bodmann CA, Koehl GE, et al: Dietary immunosuppressants do not enhance UV-induced skin carcinogenesis, and reveal discordance between p53-mutant early clones and carcinomas. Cancer Prev Res. 6:129–138. 2013. View Article : Google Scholar | |
|
Lee JH, Gaddameedhi S, Ozturk N, Ye R and Sancar A: DNA damage-specific control of cell death by cryptochrome in p53-mutant ras-transformed cells. Cancer Res. 73:785–791. 2013. View Article : Google Scholar : PubMed/NCBI |









