



Association of gut dysbiosis with first‑episode psychosis (Review)
- Authors:
- Published online on: May 24, 2024 https://doi.org/10.3892/mmr.2024.13254
- Article Number: 130
-
Copyright: © Theleritis et al. This is an open access article distributed under the terms of Creative Commons Attribution License.
Abstract
Introduction
Changes in the balance, composition and diversity of the gut microbiota (gut dysbiosis) have been found to be associated with the occurrence of various neuropsychiatric disorders and psychosis (1–3). Specifically, the gut-microbiota-brain axis, is a complex bidirectional connection system linking the gastrointestinal tract to the brain (Fig. 1), which passes through various connecting roots, such as the neuroendocrine system, the vagus nerve and the immune system (4). Additionally, the gut microbiota is responsible for the generation of short-chain fatty acids (SCFAs), neurotransmitters [dopamine, norepinephrine, norepinephrine-gamma-aminobutyric acid (GABA), glutamate and serotonin] and precursors of neurotransmitters. All these substances have been found to affect brain processes (5–7); for instance, SCFAs have been shown to prevent blood-brain barrier dysfunction and promote neurogenesis, angiogenesis and long-term memory storage (8).
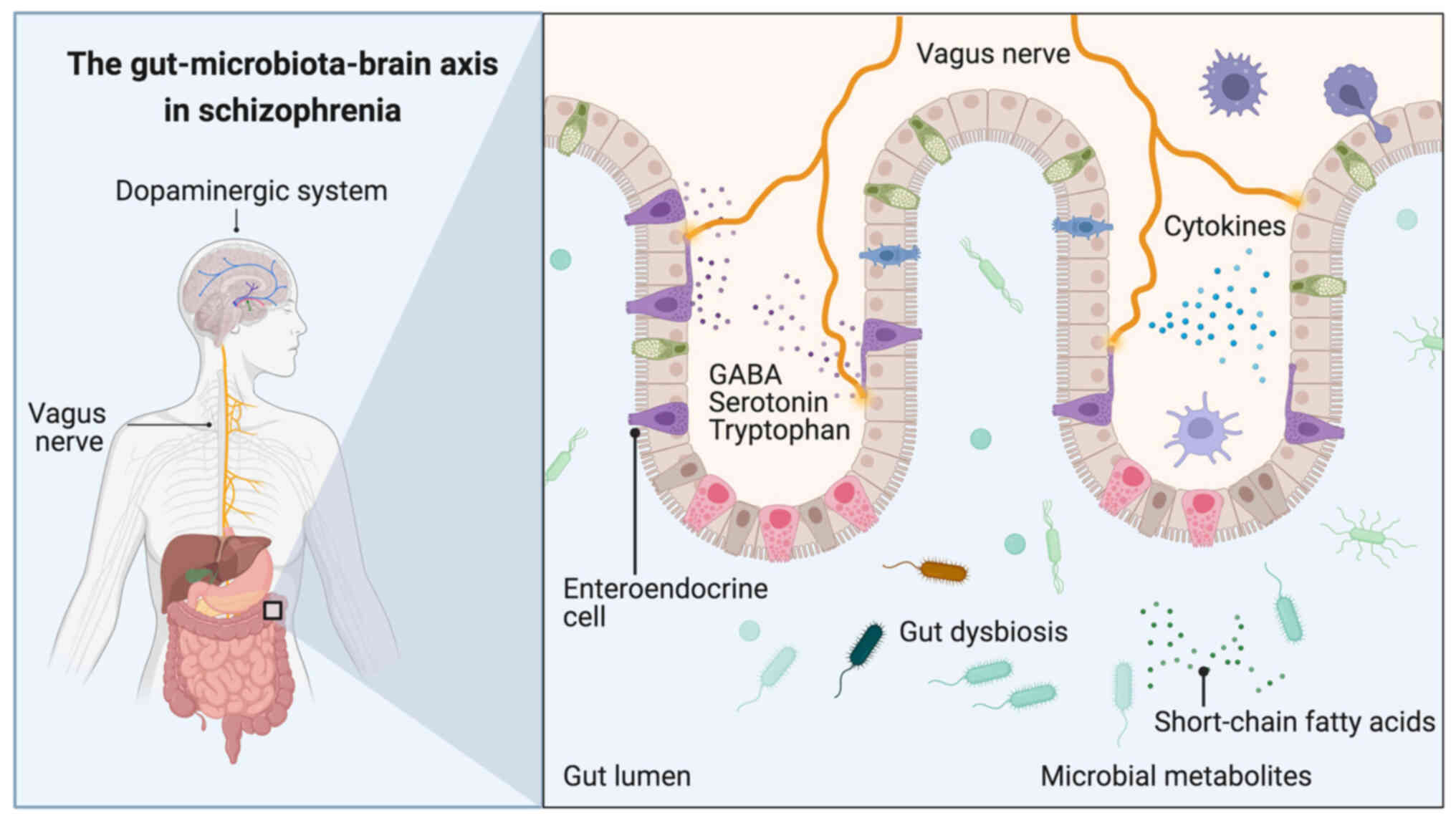
Early-life stress and various stressors encountered have been shown to be associated with gut dysbiosis, leading to irregular immunological and neuroendocrine functions, which potentially contribute to the occurrence of first-episode psychosis (FEP) (9). The treatment of psychosis could entail specific dietary interventions, along with the use of probiotics, prebiotics and fecal microbiota transplantation (10,11).
Gut microbiome and schizophrenia
The gut microbiota has been found to be associated with the occurrence of schizophrenia (SCH) (11,12). Numerous bacterial species have been found to have serotonin-synthesizing qualities (13), while abnormalities in the conversion route of tryptophan to serotonin have been reported to be linked to SCH (10). In the study by Zhu et al (14), the transfer of fecal microbiota from patients with SCH into antibiotic-treated mice resulted in psychomotor hyperactivity, diminished learning and memory, as well as in an increase in tryptophan catabolism through the kynurenine (Kyn)-kynurenic acid (Kyna) route in the peripheral and central nervous systems [Kyn and Kyna are known to influence central glutamate and serotonin systems (15)]. In the study by Zhu et al (14), the Kyn/Kyna levels were found to be associated with glutamate in the prefrontal cortex, dopamine in the striatum, and glutamate and tryptophan in the hippocampus; additionally, elevated basal levels of extracellular dopamine were found in the prefrontal cortex and 5-hydroxytryptamine in hippocampus in mice receiving SCH fecal microbiota (14).
Furthermore, it has been proposed that d-serine may be associated with SCH; consequently, changes in the gut microbiota could modulate the metabolism of d-amino acids (16,17). Moreover, Lactobacillus and Bifidobacterium in the gut may be able to produce GABA, a neurotransmitter that has been shown to be associated with SCH (16). Within this context, alterations in the glutamate glutamine-GABA cycle and reduced levels of glutamate in the brain were proposed to be associated with NMDA receptor hypofunction in SCH (18). Furthermore, the utilization of NMDA receptor antagonists (phencyclidine and ketamine) may elicit psychotic positive and negative symptoms, along with cognitive dysfunction in healthy subjects, patients with SCH and rodents (19,20). The dysregulation of dopamine in the brain has also been found to be associated with SCH. Staphylococcus can convert l-DOPA into dopamine in the human gut (21). It is noteworthy that in the study by Li et al (22), the shortage of vitamin B6 decreased dopamine levels and produced social deficits and excitation/inhibition abnormality in EphB6-deficient mice.
The levels of Saccharomyces cerevisiae and Candida albicans, markers of gastrointestinal inflammation, have been reported to be elevated in patients with SCH and FEP (23,24). The association between gastrointestinal inflammation with C-reactive protein (25) and increased gastrointestinal permeability (26) has also been reported in patients with SCH. Deficits in social behavior, a modified tryptophan-kynurenine metabolic pathway and glutamate glutamine-GABA cycles have been reported following the administration of Streptococcus vestibularis in mice, a bacterium whose levels are increased in SCH (27). As regards the differences in the gut microbiota found in subjects with SCH, the levels of the genera Succinivibrio, Megasphaera, Clostridium and Collinsella have been found to be elevated, while those of Succinivibrio have been found to be related to the severity of symptoms (28–30). In another study by Nguyen et al (31), it was observed that a plethora of Ruminococcaceae was associated with a reduced gravity of negative symptoms. At the genus level, the levels of Anaerococcus were shown to be elevated in SCH, while those of Haemophilus, Sutterella, and Clostridium were shown to be reduced; at the phylum level, the levels of Proteobacteria were found to be reduced in patients with SCH vs. controls (31). Nocera and Nasrallah (32) observed that there were associations between alterations in the gut microbiota and overall symptom gravity in SCH and negative symptomatology.
Gut microbiome and FEP
Gut dysbiosis may play a role in the occurrence of FEP and may be considered an objective for treatment (33). Below, 12 studies (presented in Table I) are discussed, which examined the effects of gut dysbiosis in treatment resistance and the psychotic symptomatology of subjects with FEP.
In the study by Schwarz et al (34), dissimilarities in the composition of the gut microbiota were reported between patients with FEP vs. the controls; Lactobacillus numbers were found to be increased and were positively associated with severity of SCH symptoms and increased positive SCH symptoms, while negative symptoms were associated with Lachnospiraceae and Ruminococcaceae. Additionally, Lactobacillus, Lachnospiraceae, Ruminococcaceae and Bacteroides spp. were shown to be negatively associated with functioning, as assessed using the Global Assessment of Functioning scale. In patients with FEP, at the family level, elevated proportions of Lactobacillaceae, Halothiobacillaceae, Brucellaceae and Micrococcineae were found, while the levels of Veillonellaceae were reduced compared with the controls; a proportion of patients with FEP with increased differences in the gut microbiota exhibited a poorer treatment efficacy following 1 year of treatment (34). In the study by He et al (35), in participants at an ultra-high risk of developing psychosis, elevated levels of Clostridiales, Lactobacillales and Bacteroidales were found, together with an increase in the levels of choline (a brain image marker of membrane dysfunction) vs. the two other study groups of high risk for psychosis and controls. Yuan et al (36) examined the changes in metabolism and gut microbiota composition following treatment with risperidone for 6 months in drug naïve subjects with FEP; the subjects exhibited abnormalities in gut microbiota composition, and had significantly reduced amounts of fecal Bifidobacterium spp., Escherichia coli, Lactobacillus spp. and elevated amounts of fecal Clostridium coccoides vs. the controls. Following 6 months of treatment, there was a significant deterioration in metabolic parameters along with significant increases in the amounts of Bifidobacterium spp. and Escherichia coli, and significant reductions in the amounts of Clostridium coccoides and Lactobacillus spp.; changes in Bifidobacterium spp. were associated with changes in weight and body mass index following 6 months of antipsychotic treatment (36). In their study, Zhang et al (37), found a significant intensification in the bacteria-fungi association network in subjects with FEP and proposed that both bacterial gut microbiota and gut microbiota promoted gut dysbiosis in subjects with SCH. In subjects with FEP, they demonstrated a plethora of harmful bacteria (Proteobacteria) and reduced levels of Faecalibacterium and Lachnospiraceae, while the gut microbiota exhibited a relative decrease in alpha diversity and an altered composition; furthermore, the subjects with FEP exhibitedan elevated amount of Chaetomium and a reduced amount of Trichoderma vs. the control group (37). In the study by Ma et al (38), it was demonstrated that subjects with psychosis, exhibited an elevation in the levels of Christensenellaceae, Enterobacteriaceae and Victivallaceae, and a reduction in the levels of Pasteurellaceae, Turicibacteraceae, Peptostreptococcaceae, Veillonellaceae and Succinivibrionaceae vs. the controls. In addition, subjects with SCH undergoing antipsychotic treatment exhibited a plethora of Peptostreptococcaceae and Veillonellaceae and the genera Megasphaera, Fusobacterium and SMB53 vs. drug-naïve subjects with FEP. A number of specific SCH-related microbiota were found to be associated with abnormal right middle frontal gyrus volume, found in subjects with SCH. In the study by Zhu et al (27), subjects with FEP demonstrated alterations in short-chain fatty acid synthesis, tryptophan metabolism and synthesis/degradation of neurotransmitters and exhibited increased alpha and beta diversity. It is worth noting that Streptococcus vestibularis, instigated deficiencies in social behavior and modified the amount of neurotransmitters in recipient mice. The authors of that study (27) emphasized the fact that microbial metabolites may be affecting the occurrence of SCH. In another study, Zhu et al (39) discovered that the beta diversity of the gut microbiota composition in subjects with FEP differed from that found in subjects with SCH and the controls. A plethora of Haemophilus was positively associated with negative SCH symptomatology, excitement, cognition and depression and an abundance of Coprococcus was negatively associated with negative SCH symptomatology (39). In the study by Yuan et al (40), following 6 months of treatment of subjects with FEP with risperidone, there was a plethora of Romboutsia and a reduced amount of Lachnoclostridium, while an increase in alpha diversity was observed. It is noteworthy that the effectiveness of treatment in patients with SCH was significantly correlated with the basal levels of Lachnoclostridium and Romboutsia (40). In the study by Li et al (41), following 6 months of treatment with risperidone, a significant increase in the serum levels of butyric acid was observed in drug-naïve subjects with FEP. The authors of that study proposed that elevated serum levels of butyric acid may be associated with the treatment efficacy in subjects with FEP, as observed with reductions in the positive and negative syndrome scale (PANSS) total and subscale scores (41). In the study by Yuan et al (42), drug-naïve subjects with FEP exhibited lower fungal alpha diversity and a significantly lower fungi-to-bacteria alpha diversity ratio vs. the controls. In addition, an elevated amount of Purpureocillium was shown to be associated with more severe SCH symptomatology and poorer cognitive ability in subjects with SCH, while an elevated amount of Aspergillus was found to be associated with an improved cognitive ability. Wang et al (43) found that an altered metabolome and dysregulated microbiome were associated with neuroactive metabolites, including GABA, tryptophan and short-chain fatty acids. They underlined the association between Ruminococcus torgues and Collinsella aerofaciens and symptom intensity, and the associations between Lactobacillus ruminis and the differential metabolites, l-2,4-diaminobutyric acid and N-acetylserotonin, and cognitive ability (43). Alterations in GABA and tryptophan neurotransmitter pathways were found to be associated with the risk of developing SCH (43). Finally, Sen et al (33), in subjects with FEP, found an increase in the amounts of Bifidobacterium adolescentis, Prevotella copri and Turicibacter sanguinis along with abnormalities in deoxyribonucleotide biosynthesis, branched-chain amino acid biosynthesis, the tricarboxylic acid cycle and fatty acid elongation and biosynthesis. Furthermore, certain gut microbes, such as Bacteroides dorei, Bifidobacterium adolescentis, Turicibacter sanguinis, Roseburia spp. and Ruminococcus lactaris were shown to be positively associated with weight gain (33).
Models explaining the link between the gut microbiome and FEP
It is worth noting that an increased intestinal permeability, chronic inflammation and oxidative stress, observed in subjects with SCH, have been proposed to be associated with changes in the microbiome (1,25). Moreover, the gut microbiota mediates the regulation of pro-inflammatory cytokines; in fact, increased amounts of IL-6, TNF-a, soluble IL-2 receptor, and elevated prostaglandin E2 levels and COX activity have been observed in subjects with FEP (44,45). Previous research using rats demonstrated that exposure to diverse early-life stressors (e.g., maternal separation or social isolation) could alter the hypothalamic-pituitary-adrenal (HPA) axis and intensify the plasma corticosterone response following acute stress, resulting in the elevated production of pro-inflammatory cytokines (46,47). Subsequently, Ko and Liu (48) observed an increase in the levels of pro-inflammatory cytokines (IL-1b, IL-6 and TNF-a) following 4 weeks of the rearing of rats in isolation. In the study by Dunphy-Doherty et al (49), it was found that continuous social isolation in rats (a confirmed animal model for SCH) resulted in changes to the gut microbiota (increases in Actinobacteria and decreases in the class Clostridia), decreased levels of hippocampal IL-6 and IL-10, and modified neurogenesis (significantly fewer BrdU/NeuN-positive cells in the dentate gyrus compared with the controls).
It has been proposed that following adversities in life (9,50), gene-environment interactions (50) and epigen-etic mechanisms (51,52) may influence the expression of genes involved in neurodevelopment, the stress response and synaptic function, which could potentially promote the occurrence of psychosis through their effects on neurotransmitters and the immune system, resulting in oxidative stress (53). Consequently, it has been suggested that genetic vulnerability and pre-/perinatal risk factors (e.g., hypoxia, maternal stress, viral infec-tion and malnutrition) may render the subject more vulnerable to environmental stressors (e.g. childhood trauma, urbanicity, migration and substance abuse) (54). Similarly, it has been suggested that early-life adversity may modify the function of the HPA axis, leading to an altered stress response (55) and heightened stress sensitivity to future stressors in adolescence and adulthood (56–58), thus promoting the occurrence of SCH symptoms through dopaminergic hyperactivity (59). Furthermore, protracted exposure to stress and to glucocorticoids may result in a decrease in hippocampal volume (60) and reduced levels of brain-derived neurotrophic factor, as observed in subjects with SCH (61–66).
Treatment of gut dysbiosis in patients with FEP
The treatment of gut dysbiosis in patients with FEP could entail specific dietary interventions, along with the use of probiotics, prebiotics and fecal microbiota transplantation. The administration of vitamin D and probiotics, including Bifidobacterium bifidum, Lactobacillus acidophilus, Lactobacillus fermentum and Lactobacillus reuteri has been shown to result in improvements in the general and total PANSS score, and in metabolic parameters, as well as in a decrease in C-reactive protein levels in patients with SCH (67). In another study, patients with SCH that were administered a probiotic with Bifidobacterium breve A-1, exhibited elevated levels of IFN-γ, IL-22, IL-1R and IL-10, and decreased TNF-α levels; these results were associated with ameliorated hospital anxiety and depression scale scores and positive and negative syndrome scale scores (68). In the study by Bravo et al (69), non-vagotomized mice fed Lactobacillus rhamnosus demonstrated decreased anxiety levels, with a modified central GABA receptor expression. In the study by Kao et al (70), the prebiotic, Bimuno galactooligosaccharide™ (B-GOS®), was administered to rats in combination with olanzapine; it decreased the acetate concentrations, blocked weight gain and improved cognitive function in psychosis. Fecal microbiota transplantation is still at the experimental stage and there are only a limited number of reports evaluating its efficacy in psychosis (71,72). Optogenetic manipulations of the gut brain axis in patients with SCH have been also proposed and are under examination (73).
Conclusions and future perspectives
The present review discussed the instrumental role of the gut microbiota in the brain and its influence on the occurrence of FEP. There is great heterogeneity observed in the changes in gut microbiota composition; however, the studies presented herein demonstrate significant GM alterations in patients with FEP vs. the controls, and underline the significant role of gut dysbiosis in the occurrence, symptom intensity and treatment efficacy in FEP. It should be noted that the composition of the gut microbiota in subjects with FEP may be influenced by various dietary, environmental and treatment factors; consequently, investigating the gut microbiota in various populations may provide the opportunity for the establishment of causal associations and the possibility for personalized treatment strategies (74–76). Within this context, further human studies are warranted, with a sufficient number of subjects who are at risk of developing psychosis, in order to further enable the evaluation of GM alterations related to FEP.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
CT and ER made substantial contributions to the conception and design of the study, and the acquisition, analysis or interpretation of data to be included in the review. CT and ER were also involved in the drafting of the manuscript, and in revising it critically for important intellectual content. MIS, MD, EA, KT, NS and DAS contributed to the design of the study. All authors have read and approved the final version of the manuscript. Data authentication is not applicable.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
DAS is the Editor-in-Chief for the journal, but had no personal involvement in the reviewing process, or any influence in terms of adjudicating on the final decision, for this article. The other authors declare that they have no competing interests.
References
|
Nguyen TT, Kosciolek T, Eyler LT, Knight R and Jeste DV: Overview and systematic review of studies of microbiome in schizophrenia and bipolar disorder. J Psychiatr Res. 99:50–61. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Van Ameringen M, Turna J, Patterson B, Pipe A, Mao RQ, Anglin R and Surette MG: The gut microbiome in psychiatry: A primer for clinicians. Depress Anxiety. 36:1004–1025. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Liu JCW, Gorbovskaya I, Hahn MK and Müller DJ: The gut microbiome in schizophrenia and the potential benefits of prebiotic and probiotic treatment. Nutrients. 13:11522021. View Article : Google Scholar : PubMed/NCBI | |
|
Gómez-Eguílaz M, Ramón-Trapero JL, Pérez-Martínez L and Blanco JR: The microbiota-gut-brain axis and its large projections. Rev Neurol. 68:111–117. 2019.(In Spanish). PubMed/NCBI | |
|
Strandwitz P: Neurotransmitter modulation by the gut microbiota. Brain Res. 1693:128–133. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Chen Y, Xu J and Chen Y: Regulation of Neurotransmitters by the gut microbiota and effects on cognition in neurological disorders. Nutrients. 13:20992021. View Article : Google Scholar : PubMed/NCBI | |
|
Stopińska K, Radziwoń-Zaleska M and Domitrz I: The microbiota gut-brain axis as a key to neuropsychiatric disorders: A mini review. J Clin Med. 10:46402021. View Article : Google Scholar : PubMed/NCBI | |
|
Michel L and Prat A: One more role for the gut: Microbiota and blood brain barrier. Ann Transl Med. 4:152016.PubMed/NCBI | |
|
Giannopoulou I, Georgiades S, Stefanou MI, Spandidos DA and Rizos E: Links between trauma and psychosis (Review). Exp Ther Med. 26:3862023. View Article : Google Scholar : PubMed/NCBI | |
|
Kang DW, Adams JB, Coleman DM, Pollard EL, Maldonado J, McDonough-Means S, Caporaso JG and Krajmalnik-Brown R: Long-term benefit of microbiota transfer therapy on autism symptoms and gut microbiota. SciRep. 9:58212019. | |
|
Rantala MJ, Luoto S, Borraz-Leon JI and Krams I: Schizophrenia: The new etiological synthesis. Neurosci Biobehav Rev. 142:1048942022. View Article : Google Scholar : PubMed/NCBI | |
|
Munawar N, Ahmad A, Anwar MA and Muhammad K: Modulation of gut microbial diversity through non-pharmaceutical approaches to treat schizophrenia. Int J Mol Sci. 23:26252022. View Article : Google Scholar : PubMed/NCBI | |
|
Roth W, Zadeh K, Vekariya R, Ge Y and Mohamadzadeh M: Tryptophan metabolism and gut brain homeostasis. Int J Mol Sci. 22:29232021. View Article : Google Scholar | |
|
Zhu F, Guo R, Wang W, Ju Y, Wang Q, Ma Q, Sun Q, Fan Y, Xie Y, Yang Z, et al: Transplantation of microbiota from drug-free patients with schizophrenia causes schizophrenia-like abnormal behaviors and dysregulated kynurenine metabolism in mice. Mol Psychiatry. 25:2905–2918. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Agus A, Planchais J and Sokol H: Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe. 23:716–724. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Ghorbani M, Rajandas H, Parimannan S, Stephen Joseph GB, Tew MM, Ramly SS, Muhamad Rasat MA and Lee SY: Understanding the role of gut microbiota in the pathogenesis of schizophrenia. Psychiatr Genet. 31:39–49. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Mikulska J, Juszczyk G, Gawrońska-Grzywacz M and Herbet M: Brain sciences HPA Axis in the pathomechanism of depression and schizophrenia: New therapeutic strategies based on its participation. Brain Sci. 11:12982021. View Article : Google Scholar : PubMed/NCBI | |
|
Zheng P, Zeng B, Liu M, Chen J, Pan J, Han Y, Liu Y, Cheng K, Zhou C, Wang H, et al: The gut microbiome from patients with schizophrenia modulates the glutamate-glutamine-GABA cycle and schizophrenia-relevant behaviors in mice. Sci Adv. 5:eaau83172019. View Article : Google Scholar : PubMed/NCBI | |
|
Krystal JH, Karper LP, Seibyl JP, Freeman GK, Delaney R, Bremner JD, Heninger GR, Bowers MB Jr and Charney DS: Subanesthetic effects of the non-competitive NMDA antagonist, ketamine, in humans: Psychotomimetic, perceptual, cognitive, and neuroendocrine responses. Arch Gen Psychiatry. 51:199–214. 1994. View Article : Google Scholar : PubMed/NCBI | |
|
Jodo E: The role of the hippocampo-prefrontal cortex system in phencyclidine-induced psychosis: A model for schizophrenia. J Physiol Paris. 107:434–440. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Luqman A, Nega M, Nguyen MT, Ebner P and Götz F: SadA: Expressing staphylococci in the human gut show increased cell adherence and internalization. Cell Rep. 22:535–545. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Li Y, Luo ZY, Hu YY, Bi YW, Yang JM, Zou WJ, Song YL, Li S, Shen T, Li SJ, et al: The gut microbiota regulates autism-like behavior by mediating vitamin B6 homeostasis in EphB6-deficient mice. Microbiome. 8:1202020. View Article : Google Scholar : PubMed/NCBI | |
|
Severance EG, Alaedini A, Yang S, Halling M, Gressitt KL, Stallings CR, Origoni AE, Vaughan C, Khushalani S, Leweke FM, et al: Gastrointestinal inflammation and associated immune activation in schizophrenia. Schizophr Res. 138:48–53. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Severance EG, Gressitt KL, Yang S, Stallings CR, Origoni AE, Vaughan C, Khushalani S, Alaedini A, Dickerson FB and Yolken RH: Seroreactive marker for inflammatory bowel disease and associations with antibodies to dietary proteins in bipolar disorder. Bipolar Disord. 16:230–240. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Severance EG, Gressitt KL, Stallings CR, Origoni AE, Khushalani S, Leweke FM, Dickerson FB and Yolken RH: Discordant patterns of bacterial translocation markers and implications for innate immune imbalances in schizophrenia. Schizophr Res. 148:130–137. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Severance EG, Yolken RH and Eaton WW: Autoimmune diseases, gastrointestinal disorders and the microbiome in schizophrenia: More than a gut feeling. Schizophr Res. 176:23–35. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Zhu F, Ju Y, Wang W, Wang Q, Guo R, Ma Q, Sun Q, Fan Y, Xie Y, Yang Z, et al: Metagenome-Wide association of gut microbiome features for schizophrenia. Nat Commun. 11:16122020. View Article : Google Scholar : PubMed/NCBI | |
|
Shen Y, Xu J, Li Z, Huang Y, Yuan Y, Wang J, Zhang M, Hu S and Liang Y: Analysis of gut microbiota diversity and auxiliary diagnosis as a biomarker in patients with schizophrenia: A cross-sectional study. Schizophr Res. 197:470–477. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Li S, Zhuo M, Huang X, Huang Y, Zho J, Xiong D, Li J, Liu Y, Pan Z, Li H, et al: Altered gut microbiota associated with symptom severity in schizophrenia. PeerJ. 8:e95742020. View Article : Google Scholar : PubMed/NCBI | |
|
Xu R, Wu B, Liang J, He F, Gu W, Li K, Luo Y, Chen J, Gao Y, Wu Z, et al: altered gut microbiota and mucosal immunity in patients with schizophrenia. Brain Behav Immun. 85:120–127. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Nguyen TT, Kosciolek T, Maldonado Y, Daly RE, Martin AS, McDonald D, Knight R and Jeste DV: Differences in gut microbiome composition between persons with chronic schizophrenia and healthy comparison subjects. Schizophr Res. 204:23–29. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Nocera A and Nasrallah HA: The association of the gut microbiota with clinical features in schizophrenia. Behav Sci (Basel). 12:892022. View Article : Google Scholar : PubMed/NCBI | |
|
Sen P, Prandovszky E, Honkanen JK, Chen O, Yolken R and Suvisaari J: Dysregulation of microbiota in patients with first-episode psychosis is associated with symptom severity and treatment response. Biol Psychiatry. 95:370–379. 2024. View Article : Google Scholar : PubMed/NCBI | |
|
Schwarz E, Maukonen J, Hyytiäinen T, Kieseppä T, Orešič M, Sabunciyan S, Mantere O, Saarela M, Yolken R and Suvisaari J: Analysis of microbiota in first episode psychosis identifies preliminary associations with symptom severity and treatment response. Schizophr Res. 192:398–403. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
He Y, Kosciolek T, Tang J, Zhou Y, Li Z, Ma X, Zhu Q, Yuan N, Yuan L, Li C, et al: Gut microbiome and magnetic resonance spectroscopy study of subjects at ultra-high risk for psychosis may support the membrane hypothesis. Eur Psychiatry. 53:37–45. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Yuan X, Zhang P, Wang Y, Liu Y, Li X, Kumar BU, Hei G, Lv L, Huang XF, Fan X and Song X: Changes in metabolism and microbiota after 24-week risperidone treatment in drug naïve, normal weight patients with first episode schizophrenia. Schizophr Res. 201:299–306. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Zhang X, Pan LY, Zhang Z, Zhou YY, Jiang HY and Ruan B: Analysis of gut mycobiota in first-episode, drug-naïve Chinese patients with schizophrenia: A pilot study. Behav Brain Res. 379:1123742020. View Article : Google Scholar : PubMed/NCBI | |
|
Ma X, Asif H, Dai L, He Y, Zheng W, Wang D, Ren H, Tang J, Li C, Jin K, et al: Alteration of the gut microbiome in first-episode drug-naïve and chronic medicated schizophrenia correlate with regional brain volumes. J Psychiatr Res. 123:136–144. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Zhu C, Zheng M, Ali U, Xia Q, Wang Z, Chenlong, Yao L, Chen Y, Yan J, Wang K, et al: Association between abundance of haemophilus in the gut microbiota and negative symptoms of schizophrenia. Front Psychiatry. 12:6859102021. View Article : Google Scholar : PubMed/NCBI | |
|
Yuan X, Wang Y, Li X, Jiang J, Kang Y, Pang L, Zhang P, Li A, Lv L, Andreassen OA, et al: Gut microbial biomarkers for the treatment response in first-episode, drug-naïve schizophrenia: A 24-week follow-up study. Transl Psychiatry. 11:4222021. View Article : Google Scholar : PubMed/NCBI | |
|
Li X, Fan X, Yuan X, Pang L, Hu S, Wang Y, Huang X and Song X: The role of butyric acid in treatment response in drug-naïve first episode schizophrenia. Front Psychiatry. 12:7246642021. View Article : Google Scholar : PubMed/NCBI | |
|
Yuan X, Li X, Kang Y, Pang L, Hei G, Zhang X, Wang S, Zhao X, Zhang S, Tao Q, et al: Gut mycobiota dysbiosis in drug-naïve, first-episode schizophrenia. Schizophr Res. 250:76–86. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Wang Z, Yuan X, Zhu Z, Pang L, Ding S, Li X, Kang Y, Hei G, Zhang L, Zhang X, et al: Multiomics analyses reveal microbiome-gut-brain crosstalk centered on aberrant gamma-aminobutyric acid and tryptophan metabolism in drug-naïve patients with first-episode schizophrenia. Schizophr Bull. 50:187–198. 2024. View Article : Google Scholar : PubMed/NCBI | |
|
García-Bueno B, Bioque M, Mac-Dowell KS, Barcones MF, Martínez-Cengotitabengoa M, Pina-Camacho L, Rodríguez-Jiménez R, Sáiz PA, Castro C, Lafuente A, et al: Pro-/anti-inflammatory dysregulation in patients with first episode of psychosis: Toward an integrative inflammatory hypothesis of schizophrenia. Schizophr Bull. 40:376–387. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
García-Bueno B, Bioque M, MacDowell KS, Santabárbara J, Martínez-Cengotitabengoa M, Moreno C, Sáiz PA, Berrocoso E, Gassó P, Fe Barcones M, et al: Pro-/antiinflammatory dysregulation in early psychosis: Results from a 1-year follow-upstudy. Int J Neuropsychopharmacol. 18:pyu0372014. View Article : Google Scholar : PubMed/NCBI | |
|
O'Mahony SM, Marchesi JR, Scully P, Codling C, Ceolho AM, Quigley EM, Cryan JF and Dinan TG: Early life stress alters behavior, immunity, and microbiota in rats: implications for irritable bowel syndrome and psychiatric illnesses. Biol Psychiatry. 65:263–267. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Plotsky PM, Thrivikraman KV, Nemeroff CB, Caldji C, Sharma S and Meaney MJ: Long-term consequences of neonatal rearing on central corticotropin releasing factor systems in adult male rat offspring. Neuropsychopharmacology. 30:2192–2204. 2005. View Article : Google Scholar : PubMed/NCBI | |
|
Ko CY and Liu YP: Isolation rearing impaired sensorimotor gating but increased pro-inflammatory cytokines and disrupted metabolic parameters in both sexes of rats. Psychoneuroendocrinology. 55:173–183. 2015. View Article : Google Scholar : PubMed/NCBI | |
|
Dunphy-Doherty F, O'Mahony SM, Peterson VL, O'Sullivan O, Crispie F, Cotter PD, Wigmore P, King MV, Cryan JF and Fone KCF: Post-weaning social isolation of rats leads to long-term disruption of the gut microbiota-immune-brain axis. Brain Behav Immun. 68:261–273. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Morgan C, Charalambides M, Hutchinson G and Murray RM: Migration, ethnicity, and psychosis: Toward a sociodevelop-mental model. Schizophr Bull. 36:655–664. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Davis EG, Humphreys KL, McEwen LM, Sacchet MD, Camacho MC, MacIsaac JL, Lin DTS, Kobor MS and Gotlib IH: Accelerated DNA methylation age in adolescent girls: Associations with elevated diurnal cortisol and reduced hippocampal volume. Transl Psychiatry. 7:e12232017. View Article : Google Scholar : PubMed/NCBI | |
|
Chen Q, Li D, Jin W, Shi Y, Li Z, Ma P, Sun J, Chen S, Li P and Lin P: Research progress on the correlation between epigenetics and schizophrenia. Front Neurosci. 15:6887272021. View Article : Google Scholar : PubMed/NCBI | |
|
Alameda L, Rodriguez V, Carr E, Aas M, Trotta G, Marino P, Vorontsova N, Herane-Vives A, Gadelrab R, Spinazzola E, et al: A systematic review on mediators between adversity and psychosis: Potential targets for treatment. Psychol Med. 50:1966–1976. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Bayer TA, Falkai P and Maier W: Genetic and non-genetic vulnerability factors in schizophrenia: The basis of the ‘two hit hypothesis.’. J Psychiatr Res. 33:543–548. 1999. View Article : Google Scholar : PubMed/NCBI | |
|
Charmandari E, Kino T, Souvatzoglou E and Chrousos GP: Pediatric stress: Hormonal mediators and human development. Horm Res. 59:161–179. 2003.PubMed/NCBI | |
|
Lardinois M, Lataster T, Mengelers R, Van Os J and Myin-Germeys I: Childhood trauma and increased stress sensitivity in psychosis. Acta Psychiatr Scand. 23:28–35. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Walker EF, Brennan PA, Esterberg M, Brasfield J, Pearce B and Compton MT: Longitudinal changes in cortisol secretion and conversion to psychosis in at-risk youth. J Abnorm Psychol. 119:401–408. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Walker EF, Trotman HD, Pearce BD, Addington J, Cadenhead KS, Cornblatt BA, Heinssen R, Mathalon DH, Perkins DO, Seidman LJ, et al: Cortisol levels and risk for psychosis: Initial findings from the North American prodrome longitudinal study. Biol Psychiatry. 74:410–417. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Sapolsky RM: Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Arch Gen Psychiatry. 57:925–935. 2000. View Article : Google Scholar : PubMed/NCBI | |
|
Vita A, De Peri L, Silenzi C and Dieci M: Brain morphology in first-episode schizophrenia: A meta-analysis of quantitative magnetic resonance imaging studies. Schizophr Res. 82:75–88. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Thompson Ray M, Weickert CS, Wyatt E and Webster MJ: Decreased BDNF, trkB-TK+ and GAD67 mRNA expression in the hippo-campus of individuals with schizophrenia and mood disorders. J Psychiatry Neurosci. 36:195–203. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Daskalakis NP, De Kloet ER, Yehuda R, Malaspina D and Kranz TM: Early life stress effects on Glucocorticoid-BDNF interplay in the hippocampus. Front Mol Neurosci. 8:682015. View Article : Google Scholar : PubMed/NCBI | |
|
Rizos EN, Rontos I, Laskos E, Arsenis G, Michalopoulou PG, Vasilopoulos D, Gournellis R and Lykouras L: Investigation of serum BDNF levels in drug-naive patients with schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 32:1308–1311. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Rizos EN, Papathanasiou M, Michalopoulou PG, Mazioti A, Douzenis A, Kastania A, Nikolaidou P, Laskos E, Vasilopoulou K and Lykouras L: Association of serum BDNF levels with hippocampal volumes in first psychotic episode drug-naive schizophrenic patients. Schizophr Res. 129:201–204. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Rizos EN, Michalopoulou PG, Siafakas N, Stefanis N, Douzenis A, Rontos I, Laskos E, Kastania A, Zoumpourlis V and Lykouras L: Association of serum brain-derived neurotrophic factor and duration of untreated psychosis in first-episode patients with schizophrenia. Neuropsychobiology. 62:87–90. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Theleritis C, Fisher HL, Shäfer I, Winters L, Stahl D, Morgan C, Dazzan P, Breedvelt J, Sambath I, Vitoratou S, et al: Brain derived Neurotropic Factor (BDNF) is associated with childhood abuse but not cognitive domains in first episode psychosis. Schizophr Res. 159:56–61. 2014. View Article : Google Scholar : PubMed/NCBI | |
|
Ghaderi A, Banafshe HR, Mirhosseini N, Moradi M, Karimi MA, Mehrzad F, Bahmani F and Asemi Z: Clinical and metabolic response to vitamin D plus Probiotic in schizophrenia patients. BMC Psychiatry. 19:772019. View Article : Google Scholar : PubMed/NCBI | |
|
Okubo R, Koga M, Katsumata N, Odamaki T, Matsuyama S, Oka M, Narita H, Hashimoto N, Kusumi I, Xiao J and Matsuoka YJ: Effect of bifidobacterium breve A-1 on anxiety and depressive symptoms in schizophrenia: A proof-of-concept study. J Affect Disord. 245:377–385. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG, Bienenstock J and Cryan JF: Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci USA. 108:16050–16055. 2011. View Article : Google Scholar : PubMed/NCBI | |
|
Kao ACC, Spitzer S, Anthony DC, Lennox B and Burnet PWJ: Prebiotic attenuation of olanzapine-induced weight gain in rats: Analysis of central and peripheral biomarkers and gut microbiota. Transl Psychiatry. 8:662018. View Article : Google Scholar : PubMed/NCBI | |
|
Hinton R: A case report looking at the effects of faecal microbiota transplantation in a patient with bipolar disorder. Aust N Z J Psychiatry. 54:649–650. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Parker G, Spoelma MJ and Rhodes N: Faecal microbiota transplantation for bipolar disorder: A detailed case study. Bipolar Disord. 24:559–563. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Patrono E, Svoboda J and Stuchlík A: Schizophrenia, the gut microbiota, and new opportunities from optogenetic manipulations of the gut-brain axis. Behav Brain Funct. 17:72021. View Article : Google Scholar : PubMed/NCBI | |
|
Nuncio-Mora L, Lanzagorta N, Nicolini H, Sarmiento E, Ortiz G, Sosa F and Genis-Mendoza AD: The role of the microbiome in first episode of psychosis. Biomedicines. 11:17702023. View Article : Google Scholar : PubMed/NCBI | |
|
Aschner M, Skalny AV, Gritsenko VA, Kartashova OL, Santamaria A, Rocha JBT, Spandidos DA, Zaitseva IP, Tsatsakis A and Tinkov AA: Role of gut microbiota in the modulation of the health effects of advanced glycation end-products (Review). Int J Mol Med. 51:442023. View Article : Google Scholar : PubMed/NCBI | |
|
Tsamakis K, Galinaki S, Alevyzakis E, Hortis I, Tsiptsios D, Kollintza E, Kympouropoulos S, Triantafyllou K, Smyrnis N and Rizos E: Gut Microbiome: A brief review on its role in schizophrenia and first episode of psychosis. Microorganisms. 10:11212022. View Article : Google Scholar : PubMed/NCBI |









